

6 курс / Клинические и лабораторные анализы / Группы крови человека
.pdfIseki, Masakia и Shibasaki [114] описали антитела, названные ими анти-Le c,
которые были получены путем иммунизации кроликов слюной Le(a −b −) секреторов и адсорбцией иммунных сывороток энзимированными эритроцитами Le(a +b −) и Le(a −b + ). Результат исследования 485 японцев сыворотками анти-Le a, анти-Le b и полученными сыворотками анти-Le c был следующим (по
Race, Sanger [197]): 107 (22,06 %) – Le(a +b −c + ), 396 (76,08 %) – Le(a −b +c −), 9 (1,86 %) – Le(a −b −c + ).
М.И. Потапов [9] получил антитела анти-Le c иммунизацией коз слюной лиц Le(a −b −) несекреторов. Полученные иммунные сыворотки были адсорбированы трипсинизированными эритроцитами Le(a + ), Le(d + ) и Le(b + ). Оставшиеся антитела реагировали с эритроцитами О(I)Le(a −b −c + ), но не О(I) Le(a −b −c −d + ).
Race и Sanger [197] сравнили специфичность козьей сыворотки, полученной М.И. Потаповым, и человеческой сыворотки анти-Le c, найденной Gunson и Latham [95]. Результаты исследования полностью совпали: эритроциты Le(a −b + ) несекреторов агглютинировались обеими сыворотками, эритроциты Le(a −b −) секреторов не агглютинировались.
Анти-Led
Эти антитела реагируют с эритроцитами Le(a −b −с −) лиц leSe / leSe и lese / leSe. Они направлены против олигосахаридов типа 1 Н, которые, так же как и другие олигосахариды Lewis, адсорбируются на эритроцитах из плазмы [109].
Антитела анти-Le d получают иммунизацией животных [7, 10, 109] слюной выделителей АВН. Аллогенные антитела анти-Le d в литературе не описаны.
Анти-Lex (анти-Leab)
У лиц Le(a −b −) находят антитела, которые по своей направленности ведут себя как анти-Le a + анти-Le b. Они могут быть простой смесью анти-Le a и антиLe b. В этом случае фракция анти-Le a легко нейтрализуется добавлением слюны донора Le(a +b −), в которой имеется субстанция Le а, но нет субстанции Le b. Адсорбированная таким образом сыворотка содержит только антитела анти-Le b. Слюну лиц Le(a −b + ) для дифференциальной нейтрализации не применяют, поскольку она содержит субстанции Le a и Le b и полностью истощает оба антитела. Антитела анти-Le a можно удалить также дифференциальной адсорбцией эритроцитами Le(a +b −).
Если антитела анти-Le a и анти-Le b представляют собой не смесь, а одно связанное антитело анти-Le ab, обозначенное анти-Le x [15, 122, 123], сепарацию антител дифференциальной нейтрализацией или адсорбцией провести не удается.
Sturgeon иArcilla [218] отметили, что антитела анти-Le x реагируют с эритроцитами Le(a +b −), Le(a −b + ) и эритроцитами новорожденных, имеющих, как известно, фенотип Le(a −b −).
Тот факт, что эритроциты детей Le(a −b −) не реагируют с моновалентными
581
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
сыворотками анти-Le a и анти-Le b, но реагируют с анти-Le x, свидетельствует о существовании на эритроцитах детей антигена Le x.
Антиген Le x присутствует также у некоторых взрослых.
Таким образом, антитела анти-Le ab (анти-Le x) не являются простой смесью антител и выявляют, помимо антигенов Le a и Le b,третий специфический анти-
ген системы Lewis – Le x (Le ab).
Andresen [16] первоначально полагал, что продукция Le x зависит от дополнительного гена Le x в системе Lewis, однако более поздние исследования, проведенные Sturgeon и Arcilla [218], позволили заключить, что продукция антигенов Le a, Le b и Le x является результатом действия одного гена Le.
В работах некоторых авторов высказываются сомнения относительно того, что Le x такой же самостоятельный антиген, как Le a и Le b, поскольку по серологической характеристике антитела анти-Le x представляют собой комбинацию специфичности анти-Le a + анти-Le b, подобно перекрестно реагирующим антителам αβ (анти-С) системы АВО.
Реакции антител анти-Le x с эритроцитами Le(a −b −) новорожденных были объяснены тем, что многие образцы пуповинной крови реагируют с сильными сыворотками анти-Le a в непрямой пробе Кумбса [62]. Иными словами, эритроциты новорожденных содержат некоторое количество вещества Le a, с которым реагируют антитела анти-Le x.
Антитела анти-Le a обычно вырабатываются секреторами субстанций АВН, антитела анти-Le b – несекреторами. Антитела анти-Le x часто, но не всегда вырабатываются Le(a −b −) секреторами. В этом их сходство с антителами анти-Le a.
Одни лица продуцируют антитела анти-Le a или анти-Le b, другие могут продуцировать оба антитела в виде раздельных фракций (анти-Le a + анти-Le b) или одной фракции анти-Le ab (анти-Le x).
ArcillaиSturgeon[21–23]показали,чтоамниотическаяжидкостьсодержитвы- сокий уровень субстанции Le a, которая проявляет себя серологически как Le x.
Антитела анти-Le x нейтрализуются слюной, содержащей Le a, слабее – слюной Le b и, что удивительно, слюной лиц Le(a −b −), являющихся несекреторами [16, 19]. Ингибиция слюной Le b (т. е. слюной секреторов) более сильная, чем слюной лиц Le(a −b −) несекреторов. Уместно напомнить, что лица Le(a −b −) могут быть секреторами и несекреторами.
То обстоятельство, что антитела анти-Le x ингибируются слюной Le(a −b −) несекреторов, подтверждает существование антигена Le x как самостоятельной единицы.
Химическая структура антигена Le x, как полагаютSchenkel-Brunner и соавт. [203, 205] и другие авторы [48, 106, 185, 240], близка детерминантам, определяемым с помощью антител анти-Le a и анти-Le b.
Для сравнения: перекрестно реагирующий антиген С в системе АВО (по Винеру) присутствует на эритроцитах А и В. Антитела анти-С представляют
582
собой несепарируемый агглютинин αβ (анти-А,В), присутствующий в сыворотках лиц О(I) наряду с сепарируемыми α и β.
Всистеме резус описаны антитела анти-DC, реагирующие с антигенами D, C
иантигеном G, который, как правило, сопровождает D и C, но иногда встречается на DC-отрицательных эритроцитах (фенотип cdeG).
Таким же перекрестно реагирующим антигеном, по-видимому, является антиген Le x в системе Lewis.
Антитела анти-Le x (как анти-Le a и анти-Le b) обычно имеют не аллоиммунное происхождение, обладают способностью связывать комплемент и могут проявлять гемолитические свойства in vitro.
Анти-А1Leb и анти-ВLeb
В 1968 г. Seaman и соавт. [206] при выполнении пробы на индивидуальную совместимость эритроцитов донора с сывороткой больного Siedler нашли антитела анти-A1Le b. Сыворотка реагировала с эритроцитами, содержащими оба антигена – A1 и Le b, но не реагировала с эритроцитами ОLe(b + ) и А1Le(b −), содержащими эти антигены порознь. Вскоре 2 такие же сыворотки были най-
дены Grookston и соавт. [61] и Gundolf [94].
Антитела анти-A1Le b нейтрализуются слюной всех секреторов А независимо от Lewis-групповой принадлежности (Grookston и соавт. [61]).
У лиц с фенотипомA1Le b антигенA1Le b присутствует в слюне.
Наряду с антителами анти-A1Le b встречаются антитела анти-ВLe b, реагирующие с антигеном Le b, когда тот присутствует на эритроцитах вместе с групповым антигеном В.
Антитела анти-А1Le b и анти-ВLe b направлены против олигосахаридов Le b типа 1, к которым добавлены имммунодоминантные сахара А или В, что и приводит к формированию антигенных детерминант, определяемых указанными антителами.
Для сравнения: антитела анти-Le bL реагируют только с эритроцитами О(I), А2Le(b + ); а анти-А1Le b и анти-ВLe b – с эритроцитами, несущими Le b и соответствующую А- или В-детерминанту.
Tilley и соавт. [228], Crookston и соавт. [90] установили, что у людей А1Le b и ВLe b в плазме присутствуют гликосфинголипиды, несущие соответственно антигены А1Le b и ВLe b. Такие лица по набору генов относятся к A1, Le, Se и B, Le, Se соответственно. Насколько известно, ни анти-А1Le b, ни анти-ВLe b не дают реакций при переливании компонентов крови (Issitt,Anstee [115]).
Анти-A1Led и анти-BLed
В 1958 г. Andresen [14], обследуя больного раком желудка с фенотипом А2Le(a −b + ), нашел антитела, которые реагировали с эритроцитами A1Le(a −b −) секреторов, менее сильно – с эритроцитами A2Le(a −b −) секреторов. Автор предположил,чтогенSe вотсутствиегенаLe специфическивидоизменяетэкспрессию
583
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
антигенаАичтообнаруженныеимантитела(известныевлитературекакMagardантитела) выявляли антиген А1Le d. Последующие исследования (Hirsch и соавт., 1975) подтвердили предположение Andresen. Magard-сыворотка была первой из обнаруженных сывороток со специфичностью анти-А1Le d. Эти антитела, как теперь известно, реагируют с олигосахаридами типа 1А (производными цепей типа 1Н). Анти-A1Le d, так же как анти-BLe d, не дают трансфузионных реакций.
Клиническое значение
Большинство антител Lewis не способно разрушать эритроциты, содержащие соответствующие антигены, in vivo [111]. Холодовые антитела анти-Le IgM, температурный оптимум которых ниже комнатной температуры 20–22 оС, не активны при температуре тела человека. Неполные антитела анти-Le IgG, хотя и являются тепловыми, однако не вызывают посттрансфузионных осложнений как истинные антиэритроцитарные антитела.
Выделяют четыре причины, по которым Lewis-несовместимость не проявляет себя клинически [115].
Во-первых, большинство антител, особенно анти-Le b, имеет низкую активность. Многие из них не реагируют с нативными эритроцитами и определяются только с помощью стандартных эритроцитов, обработанных протеолитическими ферментами (фицин, папаин, бромелин). Такие ферментзависимые антитела в реакциях несовместимости in vivo не участвуют.
Во-вторых, антигены Lewis, присутствующие в донорской плазме (если переливают цельную кровь) и в том небольшом количестве плазмы, которое имеется в эритроцитарном концентрате, нейтрализуют антитела Lewis реципиента. Взаимодействие антител с растворенными в плазме субстанциями происходит быстрее, чем с субстанциями, фиксированными на мембране эритроцитов, и деструкция эритроцитов не успевает развиться.
В-третьих, перелитые эритроциты утрачивают свои антигены Lewis в плазме реципиента. Например: эритроциты Le(a −b + ), перелитые реципиенту Le(a −b −), имеющему антитела анти-Le b, становятся Le(a −b −), а олигосахариды Le b, смытые с эритроцитов, нейтрализуются присутствующими антителами анти-Le b.
В-четвертых, если антитела имеют специфичность анти-Le bH, они способны реагировать только с эритроцитами ОLe(a −b + ), но не эритроцитами АLe(a −b + ). Таким образом, для реципиентов группы А(II), содержащих антитела анти-Le bH, любые эритроциты А(II) будут совместимыми.
Совместимость донора и реципиента по групповым антигенам, растворенным в плазме крови, в трансфузиологии не учитывают.
Иммуносерологам и трансфузиологам известно, что универсальную плазму группы AB(IV), содержащую группоспецифические субстанции АВО, переливают в большом количестве (2–2,5 л и более) реципиентам A(II) и B(III), имеющим естественные антитела против этих антигенов. Однако при этом какихлибо осложнений не наблюдают.
584
Антигены системы Gm при переливании плазмы также не учитывают, хотя некоторые реципиенты (по нашим подсчетам, около 5 %) содержат антитела против иммуноглобулинов переливаемой плазмы. Однако и в этих случаях трансфузии не сопровождаются реакциями.
По-видимому, взаимодействие растворимых антигенов с антителами не приводит к посттрансфузионным реакциям, но когда мишенью становятся нерастворимые антигены, фиксированные на эритроцитах, последние разрушаются, что проявляется клинически в виде тяжелого посттрансфузионного осложнения.
Антигены Lewis, являясь по своей природе водорастворимыми, в основной своей массе находятся в плазме и реагируют с соответствующими антителами в жидкофазной системе. Эритроциты, содержащие антигены Lewis в значительно меньшем количестве, чем плазма, остаются интактными. Это, на наш взгляд, пятая причина того, почему несовместимость донора и реципиента по системе Lewis не вызывает посттрансфузионных реакций.
В экспериментах по приживлению эритроцитов, меченных Cr51,in vivo установлено, что время циркуляции эритроцитов Le(a −b + ) у реципиентов, имеющих анти-Le b-антитела, такое же, как у реципиентов без антител. Немедленных или отсроченных трансфузионных реакций не наблюдали. Уровень гемоглобина после переливания Le b-несовместимых эритроцитов повышался как при совместимой трансфузии и оставался одинаковым в течение нескольких недель.
При огромном объеме трансфузиологической помощи в современной медицинской практике трансфузионные реакции, вызванные антителами антиLe a, все-таки регистрируют (М.А. Умнова и др. [12], Krieger, Simmons [136], Brendemoen,Aas [43] и др. [160, 164, 191]).
Jesse, Sheek [119] описали острую гемолитическую реакцию средней тяжести у 21-летней африканки, которой в связи с осложненным спонтанным выкидышем 4 раза перелили кровь. Перекрестные пробы на индивидуальную совместимость (тест с центрифугированием и непрямая реакция Кумбса) перед каждой трансфузией были отрицательными. После 4-го переливания у женщины появились боли в пояснице, озноб, температура, гематурия. Кроме анти-Le b-антител, в сыворотке женщины других антиэритроцитарных антител не обнаружено. Гемолитическая реакция была кратковременной, осложнение через 2 дня было купировано. Авторы полагали, что картина посттрансфузионного осложнения была вызвана анти-Le b-антителами. Другие случаи гемолитических посттрансфузионных реакций, обусловленных антителами антиLe b, не описаны.
Aubuchon и соавт. [26], van Loghem и соавт. (цит. по [6]) привели случаи посттрансфузионных реакций, обусловленных антителами анти-Le x. Для таких реципиентов практически все доноры несовместимы. Однако даже в этих случаях, как отмечают Waheed и соавт. [233], посттрансфузионные реакции крайне редки.
585
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
В акушерской практике зарегистрированы единичные случаи гемолитической болезни новорожденных, связанные с Lewis, которые, однако, вызывали сомнение относительно их обусловленности именно этим фактором. Mollison и соавт. [173], анализируя эти работы, не нашли достаточного клинического и гематологического подтверждения.
Как показали Spitalnik и соавт. [213], в 12 случаях из отобранных ими 13 пар мать – плод в крови матери присутствовали одновременно IgM и IgG анти-Le a-антитела, в пуповинной крови новорожденных имелись антитела анти-Le a IgG, регистрируемые только высокочувствительным ферментсвязывающим имммуносорбентным методом. Гемолитических реакций у новорожденных не бвло. Полученные данные позволили авторам заключить, что антитела анти-Le a IgM через плаценту не проходят, а антитела анти-Le a IgG легко проникают через плаценту, однако не вызывают разрушения эритроцитов плода, поскольку антигены Lewis у плодов и новорожденных отсутствуют.
Ингибиция Lewis-антител
Lewis-антитела нейтрализуют с целью идентификации антител другой специфичности, присутствующих в сыворотке. Для этого используют образцы слюны, тестированные на наличие субстанций Le a, Le b и АВН, а также вытяжки из гуммиарабика или синтетические антигены Lewis (Spitalnik и соавт., цит. по [115]). Цельная слюна нередко вызывает гемолиз эритроцитов, поэтому ее предварительно разводят раствором натрия хлорида и кипятят, чтобы устранить присутствующую в ней слизь. Методика подробно описана П.Н. Косяковым [6] и Judd [125].
Mollison и соавт. [173] описали случай, когда пациенту Le(a −b −), имеющему антитела анти-Le a + Le b, гемолизирующие эритроциты в тестах in vitro, с целью нейтрализации антител ввели парентерально очищенный концентрат Le a и Le b. Антитела пациента были нейтрализованы in vivo, после чего ему было перелито несколько доз крови Le(a −b + ) без каких-либо реакций. Через несколько дней антитела анти-Le a и анти-Le b снова появлялись в плазме пациента, теперь уже в более сильной форме, чем до нейтрализации. Прямая проба Кумбса стала положительной, поскольку перелитые ранее эритроциты были Le(a −b + ) и сенсибилизировались антителами in vivo, однако гемолиза не наблюдалось. Вскоре циркулирующие эритроциты приобрели фенотип реципиента Le(a −b −) и прямая проба Кумбса стала отрицательной.
Нейтрализация антител in vivo была применена также Andorka и соавт., Pelosi и соавт. (цит. по Issitt,Anstee [115]), но широкого распространения не получила: в значительной степени в связи с тем, что в этих модельных экспериментах даже сильные антитела, деструктирующие эритроциты in vitro, не проявляли реактогенных свойств in vivo.
586
Хромосомная локализация
По данным Mollicone и соавт. [172], Nishihara и соавт. [179], KukowskaLatallo и соавт. [137], Ball и соавт. [28], Koda и соавт. [134], Lamm и соавт. [138], генный локус Lele (FUT3) расположен на коротком плече хромосомы 19 в позиции 19р13.3.
На этой хромосоме располагаются гены, кодирующие другие фукозилтрансферазы: FUT1, FUT2, FUT5 и FUT6. Некоторые из них могут влиять на продукцию антигенов Lewis, придавая им специфические особенности. FUT6 и FUT2 участвуют в синтезе тканевого антигена Le y.
Ген le является молчащим аллелем гена Le. При генотипе lele Le- генспецифическая фукозилтрансфераза не вырабатывается.
На длинном плече хромосомы 19 располагаются также гены LW, Lutheran, Hh и Sese, ген пептидазы D (Pep D) и ген третьего компонента комплемента – C3.
Kukowska-Latallo и соавт. [137] клонировали ген Le в COS-клетках и получили рекомбинантные фукозилтрансферазы α(1→3) и α(1→4), способные синтезировать антигены Le a, Le b и X (SSEA-1). Клонированные трансферазы являлись продуктом именно Le-гена и были однотипны с Lewis-трансферазой, полученной из человеческого молока [151, 195].
Suh и соавт. [220] описали несколько мутаций в генах FUT2 и FUT3, при которых антигены Lewis не вырабатывались.
Слабый (Lew)-фенотип
Слабая выраженность антигенов Lewis обусловлена аминокислотными заменамивгенах,контролирующихпродукциюфукозилтрансфераз[71,134,172,180].
Koda и соавт. [134] нашли у японцев Le(a −b −) 2 мутации в гене FUT3 (Leu 20 →Arg, Gly 170 → Ser). Опыты с клонированием гена Le в COS-клетках показали,чтозаменаGly170→Serприводиткпродукциинеактивногофермента,тогда как при замене Leu 20 →Arg вырабатывалась активная фукозилтрансфераза, о чем свидетельствовало появление антигена Le b на поверхности COS-клеток.
Mollicone и соавт. [172] также выявили мутацию Leu 20 → Arg у индонезийцев Le(a −b −), которые содержали антигены Lewis в слюне. Такие же замены описаны у японцев [180] и шведов [71]. Замена аргинина на лицин приводила к экспрессии слабовыраженного антигена Lewis.
Nishihara и соавт. [179] и Mollicone и соавт. [172] описали мутацию Ile 356 → Lys. Гетерозиготы по этой мутации, 18 из 19 индонезийцев Le(a −b −), не содержали антигенов Lewis ни на эритроцитах, ни в слюне.
Cooling и Gu [59] исследовали с помощью ПЦР 15 афроамериканцев, имевших фенотип Lenull, Отмечены варианты мутаций в FUT3, которые приводили к продукции неактивного фермента и в результате формировали нулевой фенотип. Одну из мутаций (G →A) в нуклеотиде 13 обнаружили у 50 % обследованных (рис. 9.3), она приводила к замене Gly 5 → Ser. У других обследованных обнаружили мутации G → Т в нуклеотиде 1022, G → A в нуклеотиде 484, G →
587
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
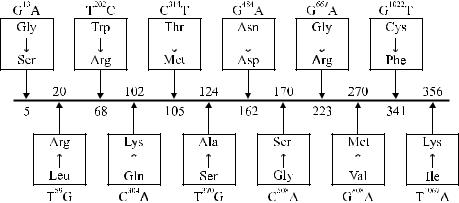
Aв нуклеотиде 667. Они приводили к замене Cys 341 → Phe, Ile 356 → Lys, которые также сочетались с низкой активностью фукозилтрансферазы.
Рис. 9.3. Мутации в гене FUT3 у лиц с фенотипом Lewisnull. Двенадцать нуклеотидных замен, приводящих к продукции неактивной фукозилтрансферазы, по Cooling, Gu [59].
Две мутации, Trp 68 → Arg и Thr 105 → Met, найдены Elmgren и соавт. [71]. Для того чтобы определить их влияние на активность фукозилтрансферазы, Elmgren и соавт. [70] сконструировали 2 химеричных протеина: FUT3 с Trp 68 →Arg и FUT3 с Thr 105 → Met. При первой замене продуцировался фермент с низкой активностью, тогда как при второй замене активность фермента соответствовала норме. Авторы пришли к выводу, что гомозиготность по Trp 68 → Arg дает фенотип Le(a −b −).
Orntoft и соавт. [187] описали мутацию (С 445 А) у человека Le(a −b −), больного раком. Эта мутация приводила к продукции метионина в позиции 146, однако такая же мутация обнаруживалась и у здоровых лиц.
Mollicone и соавт. [171] установили, что примерно 9 % жителей острова Ява не имеют α(1,3)-фукозилтрансферазы (FUT6), обычно присутствующей в плазме человека, однако содержат антигены Lewis.
При клонировании гена FUT5 от лиц с дефицитом α(1,3)-фукозилтрансферазы найдены 3 точки мутаций: Arg 173 → Cys, Pro 187 → Leu, Thr 388 → Met, кото-
рые, однако, не сказывались на активности фермента.
При клонировании гена FUT6 выявлены замены Pro 124 → Ser, Glu 247 → Lys, Tyr 315 → стоп-кодон. Две последние замены приводили к продукции неактивного фермента. Замены Glu 247 → Lys и Tur 315 → стоп-кодон в FUT6, которые сочетались с дефицитом α(1,3)-фукозилтрансферазы, обнаружены среди полинезийцев и шведов (Larson и соавт. цит. по [115]). Таким образом, ген FUT6 отвечает за активность α(1,3)-фукозилтрансферазы в плазме у людей и как другие гены FUT полиморфен.
588
Физиологическая роль
Считается, что адсорбция олигосахаридов Lewis на клетках пассивная [115], однако механизм, по-видимому, не столь прост. Транспортная функция эритроцитов изучена мало, хотя очевидно, что перенос многих биологически активных веществ – гормонов, ферментов, вирусов – это сложный микрофизиологический процесс, в котором немаловажная роль принадлежит серологически выявляемым структурам эритроцитов, в том числе Lewis.
Антигены Le а и Le x принимают участие в клеточной дифференцировке и опухолевой трансформации [75, 98] и рассматриваются как фактор, способствующий гематогенному метастазированию опухолевых клеток [97, 153, 222, 241]. Наряду с другими онкоэмбриональными антигенами антиген Le x, по мнению некоторых исследователей, может служить маркером прогрессирующей малигнизации [75, 83, 96, 99].
Le x хорошо выражен в эмбриональных тканях [83, 212] – отсюда его обо-
значение SSE-1 (Stage Specific Embryotic Antigen-1). Он максимально экспрес-
сирован на стадии морулы у мышей и, как полагают [212], играет определенную роль в преимплантации эмбриона.
Fukushi и соавт. [78], исследуя человеческие эмбрионы в разные сроки развития, установили, что Le x появляется через 40 дней с момента оплодотворения, достигает максимума к 50–70 дням, далее его экспрессия уменьшается.
У взрослых Le x присутствует на нейтрофилах, эпителии проксимальных канальцев почек [49, 76, 78] и желудочно-кишечного тракта [200].
Le а, Le x и их производные (Le b и Le y), обработанные сиалидазой, накапливаются в опухолевых клетках, которые в процессе малигнизации утрачивают нормальную дифференцировку [46, 79, 112, 200] и возвращаются к экспрессии антигена Le x, свойственного эмбриональным тканям.
Как показали серии исследований [34, 38, 131, 231], антигены Le x и Le а,обработанные сиалидазой и сульфатазой, являются лигандами адгезивных молекул, обеспечивающих хоминг лимфоцитов при остром воспалении.
Адгезия, так же как и трансэндотелиальная миграция лимфоцитов, обусловлена адгезивными молекулами, экспрессированными на поверхности эндотели-
альных клеток [30, 147, 152].
Антигены Le x, Le y (в меньшей степени Le а, Le b иLe d) обнаружены в грамотрицательныех бактериях Helicobacter pylori [24, 25, 159, 174], вызывающих хронический гастрит [237], язву желудка и двенадцатиперстной кишки [85], лимфому [190] и аденокарциному желудка [181, 182, 189].
По мнению Appelmelk и соавт. [20], патогенез Helicobacter-индуцированного гастрита включает аутоиммунный компонент и сводится к следующему. Антигенная мимикрия по Le x позволяет Helicobacter pylori, оставаясь незамеченными, нарабатыватьбольшоеколичествоантигенаLe x,ккоторомувитогевырабатываютсянейтрали- зующиеегоантителаанти-Le x.Последние,будучиперекрестнореагирующими,взаи- модействуютнетолькосHelicobacter,ноисэпителиемжелудка,провоцируягастрит.
589
Рекомендовано к покупке и изучению сайтом МедУнивер - https://meduniver.com/
Teneberg и соавт. [226] и другие авторы [167, 168] полагают, что Helicobacter pylori содержат нейтрофилактивные протеины, способствующие накоплению нейтрофилов в слизистой оболочке желудка.
Другие исследователи [37, 73, 113] связывают роль Helicobacter в этиологии гастрита с непосредственным их прилипанием к слизистой оболочке желудка с помощью адгезивных Le b-молекул, что приводит к локальному воспалению.
Молекулярная мимикрия, по мнению ряда авторов [64, 133, 215], лежит в основе заражения человека кровяным гельминтом Schistosoma mansoni, который синтезирует гликоконъюгаты, несущие антиген Le x. Анти-Le x-антитела, вырабатывающиеся у инвазированных людей, предотвращают повторное заражение [50, 65]. Однако эти антитела способны вызывать комплементзависимый цитолиз гранулоцитов, что приводит к нейтропении, наблюдаемой при шистозоматозе [209].
Список литературы
1.Абдина А.С. Группы крови у хакасов (гемотрансфузионные и этногенетические вопросы): автореф. дис. … канд. мед. наук. – М., 2000. – 19 с.
2.Аржелас Л.К. Цит. по: Косяков П.Н. Изоантигены и изоантитела человека в норме и патологии. – М.: Медицина, 1974. – С. 230.
3.Бронникова М.А. Особенности возрастной трансформации системы Левис // ХII
Междунар. конгр. по перелив. крови. – М.: Медицина, 1972. – С. 362–364.
4. Бронникова М.А., Гаркави А.С. Развитие изосерологических систем у человека в процессе эмбриогенеза // VII Междунар. конгр. антропол. и этнограф. наук. – М.: Наука, 1964. – 10 с.
5.Донсков С.И., Пискунова Т.М. Два случая Lewis-антител у молодых мужчин // Проблемы гематологии. – 2004. – № 2. – С. 41.
6.Косяков П.Н. Изоантигены системы Льюис (Lewis): Изоантигены и изоантитела человека в норме и патологии. – М.: Медицина, 1974. – С. 230.
7.Потапов M.И. Production of immuno-anti-Lewis sera in goats // Vox Sang. – 1976. – V. 30. – Р. 211–213.
8.Потапов М.И. (Potapov M.I.) Discovery of two new Lewis blood groups // 8-th Congr. Intern.Acad. Legal Med. Social Med. Opatija, Reijeka, 1970. – Р. 136.
9.Потапов М.И. Генетический аспект синтеза антигенов системы Lewis в слюнных железах // Генетика. – 1973. – Т. IX. – № 5. – С. 138–143.
10.Потапов М.И. Обнаружение антигена системы Lewis, свойственного эритроцитам выделителей группы Le(a −b −) // Пробл. гематол. – 1970. – № 11. – С. 45–49.
11.ТумановА.К.,ТомилинВ.В.Наследственныйполиморфизмизоантигеновиферментов крови в норме и патологии человека. – М., 1969. – 436 с.
12.Умнова М.А., Скачилова Н.Н., Ичаловская Т.А. и др. К вопросу о роли групповой системы Lewis в гемотрансфузионных осложнениях // Пробл. гематол. – 1975. – № 4. – С. 33–38.
13.Abe K., McKibbin J.M., Hfkomori S.I. The monoclonal antibody directed to difucosylated type 2 chain (Fucα1→2Galβ1→[Fucα1→3]GlcNAc; Y determinant) // J. Biol. Chem. –1983. – V. 258. – Р. 11793–11797.
14.Andersen J. Modifying influence of the secretor gene on the development of the ABH substance. A contribution to the conception of the Lewis group system // Vox Sang. – 1958. – V. 3. – P. 251–261.
15.AndresenP.H.Bloodgroupwithcharacteristicphenotypicalaspects//ActaPath.Microbiol. Scand. – 1948. – V. 24. – P. 616–618.
590