

2 курс / Гистология / Быков общий
.pdfСТРОЕНИЕ БЕЛОЙ ЖИРОВОЙ ТКАНИ
Белая жировая ткань состоит из долек (компактных скоплений адипоцитов), разделенных тонкими прослойками рыхлой волокнистой соединительной ткани, несущими кровеносные сосуды и нервы (рис. 11-2). Кровеносные капилляры и отдельные нервные волокна проникают внутрь долек, располагаясь в узких щелевидных пространствах между адипоцитами. Хотя адипоциты занимают основную часть объема жировой ткани, они составляют, по разным оценкам, лишь 20-60% числа ее клеток. Остальная часть приходится на клетки-предшественники адипоцитов, макрофаги, клетки сосудов и лейкоциты крови. Общее число адипоцитов в жировой ткани человека составляет 20-30х109 клеток; при ожирении оно может достигать 100х109 клеток. Химически белая жировая ткань на 60-85% представлена липидами, на 5-30% - водой и на 2-3% - белками.
Рис. 11-2. Строение белой жировой ткани. 1 - дольки (Д) жировой ткани, разделенные прослойками рыхлой волокнистой соединительной ткани (РВСТ) с кровеносными сосудами и нервами. Внутри Д между адипоцитам (АЦ) располагаются малодифференцированные клетки, макрофаги, кровеносные капилляры (КАП) и нервные волокна. 2 - зрелый адипоцит, содержащий одну крупную жировую каплю (ЖК), которая занимает большую часть объема цитоплазмы. Уплощенное ядро (Я) смещено к краю клетки вместе с тонким ободком окружающей его цитоплазмы. 3 - участок цитоплазмы адипоцита с многочисленными эндоцитозными пузырьками (ЭП) и мелкими липидными каплями (МЯК), сливающимися с основной (ОЛК), которая, окружена тонкими филаментами (Ф). Снаружи адипоцит покрыт базальной мембраной (БМ), в которую вплетаются ретикулярные волокна (РВ).
Адипоциты - крупные (диаметром от 25-50 до 150-250 мкм) клетки сферической формы, которые в жировых дольках, плотно прилегая друг к другу, нередко приобретают форму многогранников.
- 331 -
Ядро адипоцита уплощено и смещено к краю клетки вместе с тонким ободком окружающей его цитоплазмы (см. рис. 11-2). Оно содержит умеренно конденсированный хроматин.
Цитоплазма адипоцита содержит одну крупную жировую каплю, занимающую основную часть (до 95-98%) ее объема (по этой причине адииоциты белой жировой ткани называют однокапельными). Остальная часть цитоплазмы образует тончайший ободок, окружающий жировую каплю и расширяющийся до уплощенного полулуния в участке вокруг ядра, где расположена большая часть органелл адипоцита. Цитоплазма характеризуется развитой аЭПС, многочисленными пиноцитозными пузырьками, мелким комплексом Гольджи, небольшим количеством митохондрий, промежуточных филаментов. Обнаруживаются мелкие липидные капли, сливающиеся с основной, которая, по мнению одних авторов, окружена мембраной, а по данным других - тонкими (5-10 нм) филаментами.
При стандартных методах обработки гистологического материала липиды, находящиеся в жировой капле, растворяются спиртами и ксилолом, в результате чего адипоцит приобретает вид перстня или пустого пузырька с одним утолщенным краем (в области расположения ядра и основных органелл). Для выявления липидов на гистологических препаратах используются специальные методы фиксации и проводки материала, обеспечивающие их сохранение, а также окраски срезов (наиболее часто - суданом черным или суданом III).
Плазмолемма содержит многочисленные инвагинации (ямки), отражающие процессы формирования эндоцитозных пузырьков, а также соответствующие участкам слияния мембраны экзоцитозных пузырьков. Каждый адипоцит снаружи окружен базальной мембраной, в которую вплетаются ретикулярные волокна (образованы коллагеном III типа). Адипоциты обладают рецепторами нейромедиаторов (в частности, норадреналина), а также различных гормонов, которые влияют на новообразование и разрушение липидов (см. ниже).
ГИСТОФИЗИОЛОГИЯ БЕЛОЙ ЖИРОВОЙ ТКАНИ
Жиры, как трофические материалы, обладают преимуществами по сравнению с белками и углеводами - малым весом и небольшим объемом (в расчете на единицу энергии, получаемой при метаболических превращениях). Жировая ткань у среднего человека содержит 80% энергетических запасов тела. Она обеспечивает примерно 40-дневную потребность в энергии, а у лиц с ожирением - до годовой и более.
- 332 -
Ранее принятые взгляды на жировую ткань, лишь как на место сосредоточения инертных запасов жиров, накапливаемых в периоды усиленного питания и в дальнейшем используемых в качестве энергетических ресурсов в периоды голодания, в настоящее время полностью пересмотрены. Они уступили место представлениям, согласно которым жировая ткань обладает высокой метаболической активностью, а запасы ее липидов непрерывно динамично обновляются. Поддержание сравнительно постоянной массы жировой ткани обеспечивается равновесием между процессами отложения жиров (липогенеза) и их мобилизации (липолиза).
Отложение жиров в жировой ткани (липогенез)
Липиды, которые накапливаются в адипоцитах человека, представлены, главным образом (на 90-99%), триглицеридами (триацилглицеролами), т.е. являются эфирами жирных кислот и глицерина (глицерола). При температуре тела они находятся в жидком состоянии.
Источниками обновления липидов жировой ткани служат:
(1)хиломикроны (от греч. сhylos - сок и mikros - мелкий) - частицы диаметром 50-1000 нм, образующиеся в эпителиальных клетках кишки (энтероцитах) после всасывания продуктов гидролиза жиров из просвета кишки (рис. 11-3). Хиломикроны транспортируются в лимфу, оттекающую от кишки, и
вплазму крови;
(2)липопротеины очень низкой плотности (ЛОНП), которые синтезируются клетками печени (гепатоцитами) и транспортируются сывороткой крови. Они представляют собой частицы диаметром 30-90 нм и, подобно хиломикронам, состоят из центральной липидной части, окруженной оболочкой из молекул фосфолипидов, холестерина (холестерола), в которую погружены молекулы белка - аполипопротеина. Последние синтезируются в грЭПС и включаются в молекулу липопротеинов в аЭПС. Липопротеины транспортируются через комплекс Гольджи, где к ним присоединяются углеводные компоненты;
(3)триглицериды (триацилглицеролы), синтезируемые из углеводов самими адипоцитами, которые гидролизуются до свободных жирных кислот перед выделением в кровь.
Хиломикроны и ЛОНП подвергаются гидролизу с отщеплением триглицеридов (основная форма депонирования жирных кислот) в кро-
- 333 -
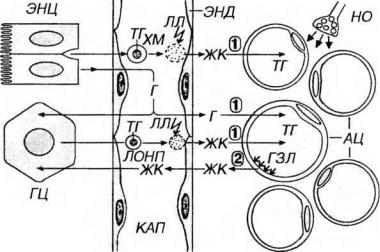
веносных капиллярах жировой ткани благодаря активности фермента липазы липопротеинов (липопротеиновой липазы). Этот фермент синтезируется адипоцитами и транспортируется в капилляры жировой ткани, где он встраивается в плазмолемму эндотелиоцитов, обращенную в просвет сосуда. Жирные кислоты, полученные при гидролизе липопротеинов, транспортируются через цитоплазму эндотелиальных клеток в межклеточное пространство, откуда захватываются адипоцитами. В цитоплазме адипоцитов (в аЭПС) жирные кислоты связываются с α-глицеро-фосфатом (процесс реэстерификации), образуя триглицериды (нейтральные жиры), которые транспортируются в жировую каплю.
Рис. 11-3. Процессы отложения (1) и мобилизации (2) жиров в жировой ткани. Энтероциты (ЭНЦ) всасывают продукты расщепления жиров из просвета кишки и ресинтезируют их с образованием триглицеридов (ТГ), которые транспортируются в лимфу, а в дальнейшем в кровь в виде хиломикронов (ХМ). ХМ расщепляются в эндотелии (ЭНД) кровеносных капилляров (КАП) ферментом липопротеиновой липазой (ЛЛ) с выделением жирных кислот (ЖК), которые переносятся в цитоплазму адипоцитов (АЦ) и после реэстерификации образуют ТГ, транспортирующиеся в жировую каплю. ТГ в АЦ могут синтезироваться из глюкозы (Г). Гепатоциты (ГЦ) захватывают ЖК из крови или синтезируют их из Г, образуя ТГ, которые транспортируются в виде липопротеинов очень низкой плотности (ЛОНП). Гидролиз ЛОНП посредством ЛЛ приводит к образованию ЖК, которые переносятся в цитоплазму АЦ, образуя ТГ. При липолизе ТГ расщепляются ферментом гормональнозависимой липазой (ГЗЛ) и в виде ЖК транспортируются в кровь. Липолиз стимулируется норадреналином, который выделяется из нервных окончаний (НО), диффундирует в пространства между АЦ и связывается с рецепторами на их плазмолемме.
- 334 -
Регуляция поглощения глюкозы жировой тканью и синтеза ею жиров из углеводов осуществляется рядом факторов, главным из которых служит инсулин. Этот гормон угнетает также выделение свободных жирных кислот жировой тканью, ингибируя активность особого фермента, осуществляющего расщепление жиров, - гормонально-зависимой липазы (липазы триглицеридов). Жировая ткань очень чувствительна к действию инсулина и является, повидимому, главной тканью-мищенью этого гормона. В отсутствие инсулина (при сахарном диабете) отмечаются повышенные уровни глюкозы, неэстерифицированных жирных кислот и липопротеинов в крови, снижение утилизации глюкозы. У таких больных главным источником энергии становятся не углеводы, а жиры.
Мобилизация жиров (липолиз)
Мобилизация жиров (липолиз) в жировой ткани осуществляется посредством нейрального и гуморального механизмов, в результате деятельности которых происходит выделение жирных кислот и глицерина в кровь. Расщепление жиров обеспечивается гормонально-зависимой липазой, которая активируется аденилатциклазой (путем образования цАМФ) при стимуляции ткани нейромедиаторами и гормонами.
Нейральная регуляция липолиза. Хотя нервные волокна,
обеспечивающие иннервацию жировой ткани, по мнению многих исследователей, являются чисто сосудодвигательными, их стимуляция приводит к липолизу. Предполагают, что норадреналин выделяется многочисленными окончаниями постганглионарных симпатических нервных волокон (входящих в состав периваскулярных нервных сплетений), распространяется по межклеточным промежуткам и связывается с рецепторами на плазмолемме адипоцитов. Норадреналин воздействует на адипоциты и как гормональный фактор, диффундируя из крови. Активность липолиза увеличивается в несколько раз в период между приемами пищи.
Гормональная регуляция липолиза. Липолиз в жировой ткани стимулируется большим числом гормонов. К липолитическим гормонам, наряду с норадреналином (обладающим у человека наиболее выраженным эффектом), относятся гипофизарные гормоны: адренокортикотропный (АКТГ), тиреотропный (ТТГ), меланоцитостимулирующий (МСГ), липотропный (ЛПГ), лютеинизирующий (ЛГ) и гормон роста (ГР). Рецепторы этих гормонов располагаются на плазмолемме адипоцитов. При взаимодействии указанных гормонов с соответствующими рецепторами происходит усиление выработки цАМФ, который в адипо-
- 335 -
цитах повышает активность гормонально-зависимой липазы. Этот фермент разлагает накопленные триглицериды на жирные кислоты и глицерин (глицерол), которые выделяются в кровоток. Дня оптимального течения липолиза требуется присутствие гормонов коркового вещества надпочечпиков (глюкокортикоидов) и щитовидной железы (тиреоидных гормонов). Чувствительность жировой ткани к липолитическим факторам с возрастом снижается.
Жировая ткань при ожирении и голодании
А. Жировая ткань при ожирении
Структурно-функциональные особенности жировой ткани при ожирении. В течение длительного времени считали, что количество адипоцитов в организме устанавливается в раннем детстве и не меняется в течение жизни. Поэтому предполагали, что увеличение массы жировой ткани при избыточном питании у взрослого человека происходит только вследствие нарастания объема (гипертрофии) адипоцитов. Этот механизм действительно характерен для большинства (примерно 80%) случаев ожирения, при которых отмечается положительная корреляция между массой тела и размерами адипоцитов. Установлено, однако, что при некоторых, наиболее тяжелых, формах ожирения, развивающихся в молодом возрасте (20% всех случаев), происходит не только гипертрофия адипоцитов, но и увеличение их числа (гиперплазия). Число адипоцитов при гиперпластической форме ожирения может увеличиваться в 3-4 раза по сравнению с таковым у индивидуумов с нормальной массой тела. Это обусловлено, вероятно, вовлечением малодифференцированных предшественников в процесс дифференцировки, завершающийся их превращением в адипоциты.
Топографические особенности отложения жировой ткани при ожирении. Увеличение массы жировой ткани при ожирении может происходить по гиноидному (от греч. gyne - женщина) или андроидному (от греч. andros - мужчина) типам (т.е. в участках нормального преобладания этой ткани в нижних или верхних частях тела - см. выше). Установлено, что гипоидное ожирение не связано с повышенным риском развития различных заболеваний. Напротив, андроидное ожирение (которое может наблюдаться и у женщин), особенно при преобладании висцерального жира над подкожным, очень часто сочетается и причинно связано с рядом тяжелых заболеваний (ишемической болезнью сердца, инфарктом миокарда, инсультом, сахарным диабетом, гипертонической болезнью).
- 336 -
Б. Жировая ткань при голодании
Структурно-функциональные особенности жировой ткани при голодании. Снижение массы тела человека в результате лечебного или вынужденного голодания сопровождается падением массы жировой ткани. Этот процесс обусловлен усилением липолиза и угнетением липогенеза, что приводит к резкому уменьшению объема адипоцитов при сохранении их общего числа в жировой ткани. Адипоциты при этом претерпевают выраженные структурно-функциональные преобразования. Единая жировая капля в их цитоплазме при мобилизации жира распадается на несколько сравнительно крупных, которые в дальнейшем становятся все более мелкими и исчезают. Адипоциты постепенно уменьшаются в размерах, из сферических становятся отростчатыми или веретеновидпыми и приобретают сходство с фибробластоподобными клетками. При возобновлении нормального питания они вновь быстро накапливают липиды и, увеличиваясь в размерах, превращаются в типичные адипоциты. Этим объясняется часто отмечаемое быстрое восстановление повышенной массы тела после отмены лечебной диеты или голодания.
Топографические особенности реакции жировой ткани на голодание.
Мобилизация жиров при голодании неодинаково затрагивает различные участки жировой ткани: в первую очередь, она отмечается в подкожных, брыжеечных и ретроперитонеальных ее скоплениях. При этом крупные адипоциты быстрее теряют липиды, чем более мелкие. Жировая ткань на ладонях, подошвах и в ретроорбитальных участках очень резистентна к процессам липолиза даже при длительном голодании. Это способствует сохранению ее опорной и пластической функции.
Репродуктивные расстройства (в особенности, у женщин)
сопровождают потерю жировой ткани при голодании (или при усиленной мышечной нагрузке, например, у спортсменок). Так, снижение массы жировой ткани, на величину, превышающую 30%, вызывает дисфункцию системы гипоталамус-гипофиз-яичники и обусловливает подавление менструального цикла и бесплодие. Предполагают, что биологический смысл этого явления (многократно описанного и у животных) заключается в выключении репродуктивной функции в отсутствие достаточных запасов питательных веществ, необходимых для обеспечения беременности и выкармливания потомства. В определенной степени, это явление может бьггь связано и с подавлением эндокринной функции жировой ткани.
- 337 -
Эндокринная функция жировой ткани
Жировая ткань вырабатывает два вида гормонов: половые стероидные гормоны (преимущественно эстрогены) и гормон, регулирующий потребление пищи - лептин.
Выработка эстрогенов. Установлено, что жировая ткань не только накапливает женские половые гормоны (эстрогены) но способна и синтезировать их благодаря высокой активности фермента ароматазы. В частности, жировая ткань служит главным источником эстрогенов у мужчин и пожилых женщин. Активность ароматазы в жировой ткани повышается с возрастом и при ожирении; она неодинакова в различных участках жировой ткани и более чем в 10 раз выше в области бедер и ягодиц по сравнению с тканью на животе и груди. Экспрессия ароматазы регулируется посредством паракринных и аутокринных механизмов, связанных с секрецией цитокинов. Способность жировой ткани активно продуцировать эстрогены может играть существенную роль и в патологии: установлено, например, что благодаря этому свойству жировая ткань, окружающая опухоль молочной железы, оказывает стимулирующее влияние на ее рост.
Выработка лептина. Баланс энергии в организме в физиологических условиях регулируется центрами, расположенными в гипоталамусе. В частности, его вентромедиальное ядро рассматривается как центр насыщения, повреждение которого вызывает ожирение. Гипоталамус контролирует активность гормональной системы регуляции потребления пищи, основным компонентом которой является пептидный гормон лептин. взаимодействующий с гипоталамическим нейропептидом Y (NPY).
Лептин (от греч. leptos - тонкий) - пептидный "фактор насыщения" (открыт в 1994 г.), вырабатываемый жировой тканью и способствующий поддержанию постоянства ее массы в организме. Он оказывает действие на уровне гипоталамуса, вызывает ощущение сытости и снижает потребление пищи.
Нейропептид Y (NPY) усиливает потребление пищи, стимулирует секрецию инсулина и накопление жира в адипоцитах. Инсулин индуцирует выделение лептина, который, в свою очередь, угнетает секрецию ИРУ и потребление пищи.
При голодании потеря массы тела (а, следовательно, и жировой ткани) вызывает снижение уровня лептина, снимая его тормозящее вли-
- 338 -
яние на гипоталамус и приводя к усиленной секреции NPY и развитию реакции на голодание. Последняя включает повышенное потребление пищи, снижение температуры тела, энергетических затрат, угнетение репродуктивной функции и усиление активности парасимпатической нервной системы. Совокупность этих изменений приводит к последующему восстановлению массы тела и жировой ткани.
Недостаточная выработка лептина или дефект лептинового рецептора в гипоталамусе быстро приводят к развитию ожирения. Последний механизм более характерен для ожирения у человека, которое чаще протекает на фоне повышенного уровня лептина, хотя описаны случаи ожирения и при сниженной продукции лептина. Уровни лептина в норме прямо пропорциональны массе тела (и содержанию жировой ткани); они несколько выше у женщин, чем у мужчин.
БУРАЯ ЖИРОВАЯ ТКАНЬ
Бурая жировая ткань содержится у человека в небольшом количестве и, в отличие от белой жировой ткани, сосредоточена лишь в нескольких четко очерченных участках тела (между лопаток, в подмышечных впадинах, на задней поверхности шеи и между ее сосудами, в воротах почек). Она сравнительно хорошо представлена у плодов человека и новорожденных (составляя у них 2- 5% массы тела). У взрослых бурая жировая ткань почти не обнаруживается, однако, полностью она, по-видимому, не исчезает. Ее содержание может даже увеличиваться у пожилых людей и при некоторых заболеваниях. Более того, она служит источником развития некоторых видов доброкачественных опухолей жировой ткани (липом). Важной особенностью бурой жировой ткани является то, что ее содержание мало меняется при недостаточном и избыточном питании.
Гистогенез бурой жировой ткани
Гистогенез бурой жировой ткани протекает сходно с развитием белой жировой ткани (см. выше). В отличие от последней, накапливающиеся в клетках липиды находятся в отдельных жировых каплях, которые сливаются друг с другом, но не образуют единой капли и поэтому не оттесняют ядро адипоцитов к периферии. В бурой жировой ткани адипоциты не достигают столь крупных размеров, как в белой.
Строение бурой жировой ткани
Бурая жировая ткань, как и белая, образована дольками, состоящими из адипоцитов бурой жировой ткани, среди которых могут на-
- 339 -
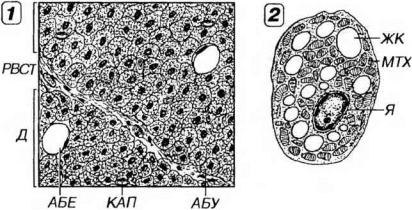
ходиться отдельные клетки белой жировой ткани (рис. 11-4). Соединительнотканные прослойки между дольками очень тонкие, а кровоснабжение долек чрезвычайно обильное. Внутри долек между адипоцитами располагаются многочисленные кровеносные капилляры и симпатические нервные волокна. Последние образуют окончания, которые, в отличие от терминалей нервных волокон в белой жировой ткани, часто плотно прижаты к поверхности адипоцитов или даже погружены в инваганированные участки их цитоплазмы. Бурый цвет ткани связан как с ее обильным кровоснабжением, так и с высоким содержанием окрашенных окислительных ферментов - цитохромов - в митохондриях адипоцитов.
Рис. 11-4. Строение бурой жировой ткани. 1 - дольки (Д), состоящие из адипоцитов бурой жировой ткани (АБУ), с отдельными клетками белой жировой ткани (АБЕ), разделенные тонкими прослойками рыхлой волокнистой соединительной ткани (РВСТ). Внутри Д между АБУ - многочисленные кровеносные капилляры (КАП) и нервные волокна. 2 - ультраструктурная организация адипоцита бурой жировой ткани. Ядро (Я) не смещено к периферии клетки; цитоплазма содержит множественные жировые капли (ЖК) и большое количество митохондрий (МТХ).
Адипоциты бурой жировой ткани существенно отличаются от аналогичных клеток в белой жировой ткани. Они имеют более мелкие размеры (до 60 мкм) и полигональную форму. Их округлое ядро располагается в центре клетки или эксцентрично (в последнем случае оно не смещено к периферии адипоцита, а цитоплазма содержит множественные жировые капли различных размеров (наиболее крупные из них достигают 25 мкм). По этой причине такие клетки называют многокапельными адипоцитами (см. рис. 11-4). В цитоплазме располагается
- 340 -