

 препятствуют образованию водородных связей
препятствуют образованию водородных связей
|
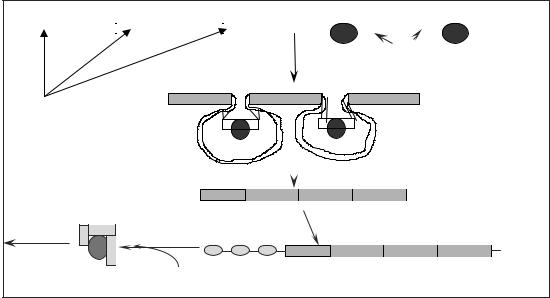
Элонгация. К 3’-ОН группе |
|
5’ |
3' |
||||
праймера |
присоединяется |
ДНК- |
|
|
Фрагменты |
|||
полимераза-III и осуществляет |
|
|
||||||
|
|
Оказаки |
||||||
синтез дочерней цепи ДНК в |
Лидирующая |
праймеры |
||||||
направлении |
5’→3’. |
Фермент не |
|
цепь |
запаздываю |
|||
способен |
синтезировать |
дочер- |
|
|
щей цепи. |
|||
|
|
|
||||||
нюю цепь ДНК на «пустом» ме- |
|
Репликационная |
||||||
сте. Эта особенность обусловлена |
|
|||||||
|
вилка. |
|
||||||
тем, что перед присоединением |
|
|
|
|||||
очередного |
|
комплементарного |
|
|
|
|||
нуклеотида ДНК-полимераза-III |
3’ |
Праймер лидирующей |
||||||
проверяет правильность встраива- |
|
цепи |
3’ 5’ |
|||||
ния |
предыдущего. |
Поскольку |
|
|
|
|||
предыдущий |
нуклеотид |
отсут- |
|
|
|
|||
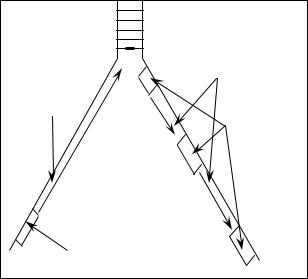
ствует – то и последующий не может быть присоединен. Поэтому для |
||||||||
работы ДНК-полимеразы-III необходим праймер (затравка). Если |
||||||||
направление синтеза дочерней цепи ДНК и направление движения ре- |
||||||||
пликационной вилки совпадают, то цепь синтезируется непрерывно и |
||||||||
называется лидирующей. Напротив, если направление синтеза ДНК и |
||||||||
движения репликационной вилки не совпадают – цепь синтезируется |
||||||||
фрагментами и называется запаздывающей. Фрагменты, образующиеся |
||||||||
в запаздывающей цепи, называют фрагментами Оказаки. Образование |
||||||||
фрагментов Оказаки обусловлено необходимостью синтеза праймера |
||||||||
каждый раз по мере продвижения репликационной вилки и обеспечи- |
||||||||
вает так же увеличение скорости репликации ДНК. |
|
|||||||
|
Праймер необходим лишь для присоединения ДНК-полимеразы- |
|||||||
III. После завершения синтеза фрагмента Оказаки праймер становится |
||||||||
не нужным. Он опознается ДНК-полимеразой-I и последовательно, |
||||||||
нуклеотид за нуклеотидом, удаляется. Вместо удаленных рибонуклео- |
||||||||
тидов встраиваются комплементарные материнской цепи ДНК дезок- |
||||||||
сирибонуклеотиды. При этом для присоединения ДНК-полимеразы-I |
||||||||
используется 3’-ОН группа предыдущего фрагмента Оказаки (или ли- |
||||||||
дирующей цепи). Сшивание синтезированных фрагментов ДНК в одну |
||||||||
цепь осуществляется ферментом ДНК-лигазой. |
|
|||||||
|
Терминация. На данной стадии происходит слияние всех репли- |
|||||||
кационных пузырей, молекулы дочерней цепи ДНК сшиваются ДНК- |
||||||||
лигазой, матрица исчерпывается, и процесс репликации прекращается. |
||||||||
|
У эукариот вместо ферментов ДНК-полимераза-I и III обнаруже- |
|||||||
ны ДНК-полимеразы α, β, γ и σ. ДНК-полимераза α реплицирует ДНК |
||||||||
в ядре клетки, β – необходима для репарации ДНК, γ – реплицирует |
||||||||
кольцевую ДНК митохондрий, σ – похожа на α, однако, функция не |
||||||||
известна. |
|
|
|
|
|
|
|
|
|
|
|
|
|
293 |
|
|
|
 информативных участков
информативных участков