

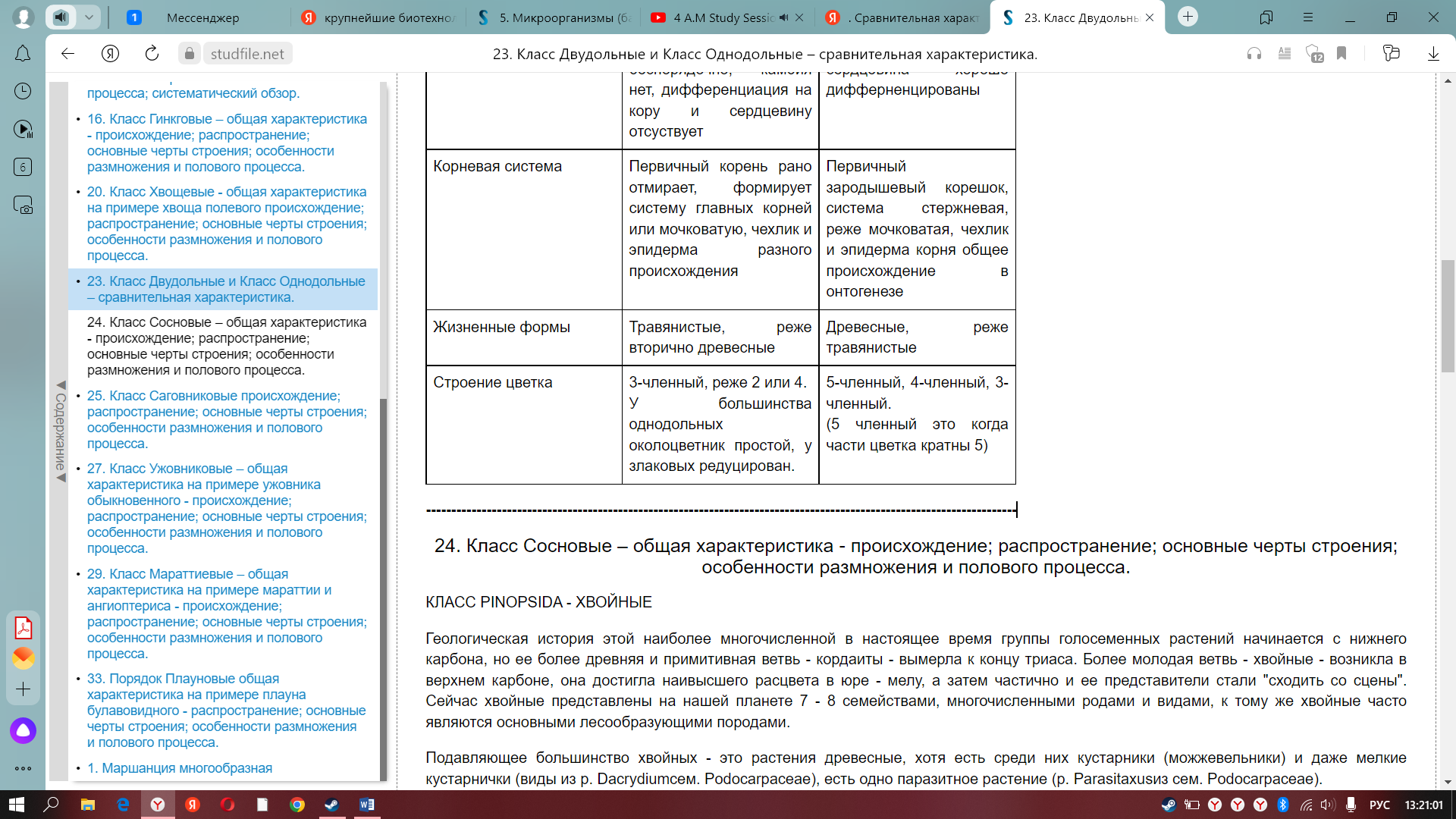
8. Сравнительная характеристика классов Двудольные и Однодольные; количественные соотношения важнейших таксонов (подклассов, порядков, семейств, родов и видов).

Отдел цветковые делят на 2 класса — двудольные и однодольные, класс двудольные — на 8 подклассов, а однодольные - на 4.
Отдел. Цветковые, или покрытосеменные
Класс I. Двудольные Подкласс 1. Магнолиевые Подкласс 2. Лютиковые Подкласс 3. Гвоздичные Подкласс 4. Гамамелисовые Подкласс 5. Диллениевые Подкласс 6. Розоцветные Подкласс 7. Губоцветные Подкласс 8. Сложноцветные |
Класс П. Однодольные Подкласс 9. Частуховые Подкласс 10. Триурисовые Подкласс 11. Лилейные Подкласс 12. Арековые |
Согласно А. Л. Тахтаджяну (1987), класс Двудольных (Dicotyledones) включает в себя 8 подклассов, состоящих из 429 семейств, приблизительно 10 000 родов и не менее 190 000 видов.
Класс подразделяется на 6 подклассов и включает 37 порядков, 122 семейства, примерно 3 100 родов и около 63 000 видов.
9. Аутэкологический и синэкологический оптимум
Оптимум вида (optimum of species) - набор условий среды, который обеспечивает виду максимальную биологическую продуктивность. Различают аутэкологический оптимум вида и синэкологический. В первом случае под оптимумом вида понимают тот набор условий среды (т.е. точку в гиперпространстве экологических факторов среды), когда вид достигает максимального развития при отсутствии конкуренции (Миркин, Розенберг, 1978). Во втором случае - аналогичный набор факторов, но при наличии конкуренции. Различия аутэкологического и синэкологического оптимума вида объясняют многие феномены природы: двувершинные распределения видов по градиенту, различная реакция на удобрения луговых видов в травосмеси и в чистом посеве (Работнов, 1983).
От оптимума вида, опосредствуемого как оптимум популяции, следует отличать оптимум индивидуума, который может не совпадать с оптимумом вида. Так, в опытах с травосмесями оптимум популяции, отражаемый максимальной долей участия вида в сообществе, наступает раньше, чем оптимум индивидуума, который опознается по максимальной величине растений
Аут- и синэкологические оптимумы и амплитуды обычно не совпадают (рис. 4), и причины этого могут быть различными. Так, оптимальные для вида значения какого-либо фактора могут различаться в зависимости от выраженности иных экологических факторов. Существенно влияет на синэкологическую амплитуду и фитоценотическая обстановка. Сильные конкуренты и виды, сильно изменяющие среду, могут вытеснять более слабые виды из благоприятных для них условий местообитания, вследствие чего их синэкологическая амплитуда становится заметно уже аутэкологической (рис. 4а). Может также происходить и сдвиг синэкологического оптимума по отношению к аутэкологическому
10. Класс Антоцеротоподобные (Anthocerotopsida). Общая характеристика
Наиболее характерные особенности следующие: своеобразный, примитивный гаметофит и сложно организованный спорофит.
Характерны пластинчатые талломы большей частью розетковидной формы диаметром 0.5-3 см, темно-зеленые. По краям талломов – меристематические клетки, которые формируют лопасти, налегающие друг на друга и придающие таллому курчавость (надрезанно-лопастные по виду).
В середине они плотно прижаты к субстрату и состоят из 6 – 10 слоев клеток, по краям же однослойные и часто загнуты вверх. К земле талломы прикрепляются при помощи простых ризоидов. Нарастают талломы при помощи верхушечных клеток, расположенных между лопастями слоевища
Анатомическая структура таллома представлена однородными паренхимными клетками. Эпидермис не выражен. В клетках таллома, как и у водорослей, имеются пластинчатые хроматофоры, внутри которых находится пиреноид. В талломе имеются слизевые полости, открывающиеся наружу. Слизь защищает таллом от высыхания. На внутренних стенках полостей имеются устьица. Внутри клеток таллома, как апланоспоры у водорослей, образуются выводковые почки. Они служат для вегетативного размножения. Образование выводковых почек связано с отделением некоторых клеток от стенок таллома. Благодаря ряду последовательных делений формируется небольшое тельце, освобождающееся по ослизнении стенок произведшей их клетки. Освободившись от материнского таллома, эти тельца разрастаются и дают новые дочерние талломы.
У многих видов образуются межклеточные полости на нижней стороне таллома, открывающиеся наружу щелевидным отверстием. Полости заполнены слизью, которая активно поглощает влагу. В них врастают нити цианобактерии рода Носток (Nostoc), являющейся симбионтом с антоцеротовидными. Колонии ностока, нарастая, увеличиваются в размерах. Вместе с ростом колонии разрастаются и полости. Цианобактерии фиксируют атмосферный азот и снабжают им свои растения-хозяева.
Большинство антоцеротовидных являются однолетними растениями, некоторые – многолетними. У однолетников таллом разрушается после созревания и рассеивания спор.
Жизненный цикл:
Антеридии и архегонии образуются на одном и том же растении и погружены в ткань таллома. Антеридии развиваются в особых полостях по одному или группами. При созревании антеридиев крыша полости разрушается, антеридии вскрываются и двужгутиковые сперматозоиды выплывают наружу.
Архегонии не имеют собственной стенки и состоят из четырехклеточной шейки, брюшной клетки и яйцеклетки. Сверху архегонии одеты поверхностными клетками таллома, которые расступаются ко времени созревания архегониия.
Канальцевые клетки расплываются в слизь, по которой сперматозоид проникает к яйцеклетке. Из оплодотворенной яйцеклетки развивается спорогоний. Зрелый спорогоний имеет вид толстой щетинки, 2 – 3 см длиной. В нижней части спорогоний расширяется в гаусторию, внедряющуюся в ткань таллома. Формирование и рост щетинки происходит за счет меристематической ткани.
У некоторых видов антоцероса спорогоний может разрастаться до 16 см. При этом он внедряется в почву, добывая оттуда воду и минеральные вещества. Спорогоний вскрывается в верхней части двумя продольными створками, обнажая при этом колонку, состоящую из бесплодной ткани. Между колонкой и створками расположены споры и пружинкообразные разветвленные клетки. Стенки коробочки сверху покрыты эпидермой с типичными устьицами, а под эпидермой располагается фотосинтезирующая ткань. При прорастании споры образуется тонкая пластинчатая протонема, на которой образуется множество точек роста. Нарастая, они формируют лопастной курчавый таллом антоцероса.
Наиболее многочисленной по видовому составу группой антоцеротовых является род Anthoceros, насчитывающий около 200 видов, распространенных в умеренных областях обоих полушарий, а также в тропиках.
11. Различные взгляды на происхождение эволюции отдела покрытосеменные ( место, время возникновения, полифилия, причины быстрораспространения и др.) Основные направления морфологической эволюции покрытосеменных.
Местом возникновения покрытосеменных со времен Дарвина считали высокие широты, согласующееся с широко распространенным тогда представлением о примитивности сережкоцветных и подкрепленное находкой остатков покрытосеменных в меловых отложениях Западной Гренландии. Впоследствии выяснилось, что гренландские покрытосеменные происходят главным образом из верхнемеловых отложений. Теперь уже довольно обширные палеонтологические материалы показывают, что первое появление покрытосеменных приходится на альб, большей частью его средние и верхние горизонты, т.е. они появились здесь примерно на 10—15 млн лет позднее, чем в низких и средних широтах.
Одно из теоретических возражений против горной гипотезы заключалось в том, что растительность склонов гор (если не говорить об альпийских лугах, аналогов которых в мезозое, вероятно, не существовало) представляет собой динамическую систему поясов, гипсометрическое положение которых не остается постоянным, смещаясь вверх или вниз под воздействием климатических изменений и других факторов. Нисходящие миграции приближают склоновые пояса к областям осадконакопления, в результате чего резко возрастает вероятность захоронения соответствующих остатков. Хорошо известны примеры нисходящих миграций в мезозое и кайнозое. Крайне маловероятно, чтобы цветковые в течение нескольких геологических периодов оставались в верхних поясах, несмотря на неоднократные изменения климата и перестройки растительности.
Знаменитый в истории Чарльз Дарвин задавался вопросом - почему так резко, буквально за короткое время, появилось огромное количество покрытосеменных (цветковых) растений и что на это повлияло. Он посвящал много работ исследованиям биологии; одно время отдавал достаточное пристрастие к растениям вместе с коллегами-ботаниками для разрешения загадки цветов в эволюции, или, как он называл «страшной тайны», ведь это противоречило взгляду с точки зрения эволюции и естественного отбора.
У каждого вида покрытосеменных растений существует своя уникальная история эволюции. Это зависит от того, какие насекомые опыляют растение. Но существует одно семейство цветковых растений, которое довело до совершенства взаимоотношения между насекомыми и цветами - это орхидеи. Благодаря уникальным взаимоотношениям, орхидеи стало огромное разнообразие, их насчитывают в мире всего 25 000 видов. Орхидеи особенно восхищали и занимали Ч.Дарвина: «Меня особенно сильно заинтересовал катасетум (вид орхидей), как впрочем, и другие экзотические орхидеи. Орхидеи интересуют меня больше всего остального в жизни» 1862г
Результаты данных обстоятельств можно увидеть в самых первых цветах в истории, а именно - семейство кувшинковых. Разнообразие ярких и разноцветных цветов, которые видоизменились из листьев, привлекали насекомых. Яркие цвета говорили о том, что в них есть очень питательная пыльца. Благодаря тому, что форма цветка образует своего рода простейшую ловушку, насекомые спотыкались и натыкались на органы полового размножения, тем самым, случайно разнося пыльцу с одного цветка на другой. Таким образом, растения заставляли обманом переносить пыльцу на другие цветы.

Покрытосемянность несомненно представляла большое преимущество в смысле защиты семяпочек и развивающихся семян от всяких неблагоприятных внешних воздействий и в первую очередь от сухости воздуха. Но одной покрытосемянностью все же трудно объяснить быстрое мощное развитие покрытосеменных и вытеснение ими господствовавших ранее на земле архегониальных растений. Русский ботаник М. И. Голенкин высказал (в 1927г) интересную гиппотезу о причинах победы покрытосеменных в борьбе за существование. Он предполагает, что в середине мелового периода по каким-то общим космогоническим причинам по всей Земле произошло резкпридаточныхое изменение освещения и влажности воздуха. Густые облака, окутовавшие ранее постоянно Землю, рассеялись и дали доступ ярким солнечным лучам, в связи с чем резко увеличилась сухость воздуха. Громадное большинство высших архегониальных растений того времени, не приспособленных и не сумевших приспособиться к яркому освещению и сухости воздуха, начало вымирать или резко сократило области своего распространения (кроме хвойных, наиболее ксерофитных).