

24. Молекулярные механизмы действия гормонов разной химической природы на клетки-мишени.
Конечными эффектами действия гормонов на клеточном уровне могут быть изменения метаболизма, проницаемости мембраны для различных ве ществ (ионов, глюкозы и др.), процессов роста, дифференцировки и деления клеток, сократительной или секреторной активности и др. Реализация этих эффектов начинается со связывания гормона со специфическими клеточны ми белками-рецепт орами : мембранными или внутриклеточными (цито плазматическими и ядерными). Эффект действия гормонов через мембранные рецепторы проявляется относительно быстро (в течение нескольких минут), а через внутриклеточные рецепторы - медленно (от получаса и более).
Действие через м ем бранны е рецепторы типично для белково-пептид ных гормонов и производных аминокислот. Эти гормоны (за исключением тироидных), являются гидрофильными и не могут проникать сквозь били пидный слой плазмалеммы. Поэтому, гормональный сигнал передается в клетку по относительно длинной цепи, которая в общем случае выглядит так: гормон —> мембранный рецептор —> мембранный фермент —> вторичный посредник —> протеинкиназа —> внутриклеточные функциональные белки —> физиологический эффект.
Соответственно действие гормона через мембранные рецепторы реа лизуется в несколько этапов:
1 ) взаимодействие гормона с мембранным рецептором приводит к изменению конформации рецептора и его активированию;
2) рецептор активирует (реже - ингибирует) связанный с ним мем бранный фермент ;
3) фермент изменяет концентрацию в цитоплазме того или иного низкомолекулярного вещества - вторичного посредника',
4) вторичный посредник активирует определенную цитоплазматичес кую протеинкиназу - фермент, катализирующий фосфорилирование и из менение функциональных свойств белков;
5) протеинкиназа изменяет активность внутриклеточных функцио нальных белков, регулирующих внутриклеточные процессы (ферментов, ионных каналов, сократительных белков и т.п.), в результате чего реализу ется тот или иной конечный эффект действия гормона, например, ускоре ние синтеза или распада гликогена, запуск мышечного сокращения и др.
В настоящее время известны четыре типа ферментов, связанных с мембранными рецепторами гормонов, и пять основных вторичных посредников.
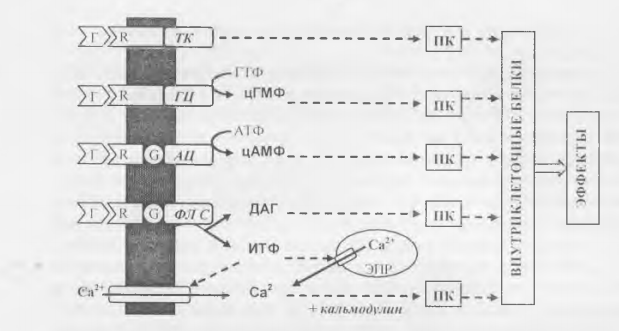
Обозначения: Г - гормоны; R — мембранные рецепторы; G - G-белки; Ж - тирозин-киназа; Г Ц - гуанилатциклаза; А Ц ~ аденилатциклаза; Ф.П С - фосфолипаза С; фл - мембранные фосфолипиды; ИТФ - инозитолтрифосфат, Д АТ - диацилглицерол; ЭПР - эндоплазматический ретикулум; ПК - различные протеинкиназы.
25. Нейросекреторная функция гипоталамуса. Рилизинг-факторы, их характеристика. Гипоталамо-гипофизарные связи.
Особое значение нейросекреторные элементы имеют в осуществлении защитных и приспособительных реакций организма.
В супраоптическом и паравентрикулярном ядрах располагаются клеточные тела, отростки которых образуют гипоталамо-гипофизарный тракт, идущий в заднюю долю гипофиза. Последняя выступает в роли гемального органа, где наполненные секретом аксонные терминали функционально взаимодействуют с выносящими секрет сосудами. Клетки супраоптического и паравентрикулярных ядер получают синаптический вход от разных структур мозга. Это типичные нейроны.
Они генерируют потенциалы действия, распространяющиеся по аксону. Достигая аксонной терминали, потенциал вызывает секрецию пептидного гормона, которая осуществляется по механизму экзоцитоза, т. е. того самого механизма, с помощью которого выделяются и медиаторы. По своей ультраструктуре эти клетки отличаются от других нейронов наличием крупных (до 190 нм) электронно-плотных секреторных гранул, которые окружены специальной мембраной. Особенно много гранул в аксональных терминалях.
В области секреторной мембраны аксона локализуются мелкие пузырьки, не имеющие плотного содержимого. Они являются, по-видимому, либо продуктами пиноцитоза, либо остатками изливших содержимое секреторных гранул.
Секретируемый нервными клетками материал перемещается по аксонам гипофизарно-гипоталамического тракта в форме коллоидных гранул. Перерезка тракта сопровождается накоплением нейросекрета в его проксимальном конце и истощением в дистальном. Различного рода стрессорные воздействия, жажда, обезвоживание также ведут к истощению нейросекрета. У крыс, например, стресс вызывает истощение запаса нейросекрета в течение 10 мин.
Не менее важна роль гипоталамуса и в секреции аденогипофизарных гормонов. В состав мелкоклеточных ядер вентральной гипоталамической области входят клетки, которые вырабатывают нейрогормоны, регулирующие секреторную активность тройных желез аденогипофиза.
Рилизинг-факторы, их характеристика.
Риллизинг-факторы-класс пептидных гормонов гипоталламуса, общим свойством которых является реализафия их эффектовчерез стимуляцию синтеза и секреции в кровь тех или иных тропных гормонов передней доли гипофиза. Подразделяются на 2 группы:
1)Либерины- гормоны белковой природы. Вырабатываются в гипоталамусе, далее поступают в гипофиз и активируют синтез и выделение в кровь гормонов передней доли гипофиза. Основными либеринами являются соматолиберины, кортиколиберины, тиреолиберины и т.д.
2) Статины- гормоны белковой природы. Вырабатываются в гипоталамусе, далее поступают в гипофиз и подавляют синтез и выделение в кровь гормонов передней доли гипофиза. Основными статинами являются соматостатины, кортикостатины, тиреостатины и т.д.
Гипоталамо-гипофизарные связи.
Гипоталамо-гипофизарная система — объединение структур гипофиза и гипоталамуса, выполняющее функции, как нервной системы, так и эндокринной. Этот нейроэндокринный комплекс является примером того, насколько тесно связаны в организме млекопитающих нервный и гуморальный способы регуляции.
Строение: Гипоталамо-гипофизарная система состоит из ножки гипофиза, начинающейся в вентромедиальной области гипоталамуса, и трёх долей гипофиза: аденогипофиз (передняя доля), нейрогипофиз (задняя доля) и вставочная доля гипофиза. Работа всех трёх долей управляется гипоталамусом с помощью особых нейросекреторных клеток. Эти клетки выделяют специальные гормоны — рилизинг-гормоны. Релизинг-факторы попадают в гипофиз, а точнее в аденогипофиз через воротную вену гипофиза.
Гормоны гипоталамо-гипофизарной системы:
Под влиянием того или иного типа воздействия гипоталамуса, доли гипофиза выделяют различные гормоны, управляющие работой почти всей эндокринной системы человека. Исключение составляет поджелудочная железа и мозговая часть надпочечников. У них есть своя собственная система регуляции.