

Биофизика и биохимия фотосинтеза
.pdfНаукові матеріали
Ботаніка, фізіологія рослин Агрохімія, ґрунтознавство Хвороби та шкідники Захист рослин Прогнозування Програмне забезпечення Селекція та генетика Насінництво Зберігання та переробка Економіка та логістика Екологія Ентомологія
Головна / Ботаніка, фізіологія рослин
Биофизика и биохимия фотосинтеза
источник: Физиология растений - 3-е изд., М: 1988 г. // Лебедев С.И.
Согласно суммарному уравнению, основными продуктами фотосинтеза являются углеводы. Фотосинтез также служит источником свободного кислорода на нашей планете. Элементарный химический состав растений может быть представлен следующими средними данными, %: С — 45, О — 42, Н — 6,5, N — 1,5, зола — 5. Таким образом, в растении на долю углерода и кислорода приходится наибольшее количество. Установлено, что 95% своей массы растение строит за счет углекислого газа, который получает из воздуха, и воды.
Углерод поступает в растение в процессе фотосинтеза. Например, как показали расчеты, для создания урожая зерна озимой пшеницы 4 т с 1 га растения поглощают из почвы около 150 кг азота и 300-500 кг других элементов минерального питания (фосфор, сера, калий, кальций, кремний и микроэлементы), при этом они должны усвоить не менее 4500 кг углерода и подвергнуть разложению почти 16 т воды, из которой примерно 2 т водорода используется для восстановления углекислого газа, а 14 т свободного кислорода выделяется в атмосферу. Чтобы накопить в процессе фотосинтеза указанное количество углерода, растения на 1 га должны на протяжении вегетационного периода усвоить 20-25 т углекислого газа, т. е. 150-300 кг в день. Для такой грандиозной химической работы растения пшеницы поглощают 16,72 млрд кДж солнечной радиации, из них 9,2 млрд кДж составляет фотосинтетически активная радиация (свет с длиной волны 380-710 нм). В органической массе урожая растения запасают около 167,2 млн кДж энергии. Остальная энергия превращается в тепло и расходуется на испарение воды (транспирацию).
К процессу фотосинтеза применимы три основных правила (принципа) фотохимии: первое — химическое изменение может производить только поглощенный свет (закон Гротгуса — Дрейпера); каждый поглощенный фотон активирует только одну молекулу (закон Штарка — Эйнштейна); третье — вся энергия кванта сообщается при поглощении света одномуединственному электрону, вследствие чего он поднимается на более высокий энергетический уровень (постулат Эйнштейна). Таким образом, фотохимический эффект прямо пропорционален количеству воспринятой веществом энергии. Известно, что электрон представляет заряженную частицу, которая движется по некоторой орбите вокруг ядра атома. Энергия электрона зависит как от положения орбиты в пространстве, так и от скорости его движения по орбите. Увеличение энергии электрона в результате поглощения фотона света может быть использовано для переноса его на орбиту, обладающую более высокой энергией, чем исходная, либо для увеличения скорости движения электрона вокруг ядра по сравнению с той, с какой он двигался до поглощения света.
Современные научные представления о фотохимических реакциях, входящих в процесс фотосинтеза, основываются на квантовой теории света. Согласно этой теории, каждая молекула вещества при фотохимических реакциях, т. е. при превращении вещества под влиянием света, поглощает частицу его — фотон, или квант, который имеет достаточный запас энергии для того, чтобы вызвать в молекуле соответствующие изменения. Основное действие света определяется энергией как отдельных фотонов, так и световых потоков.
Если квант меньше, чем необходимо энергии для данной реакции, то превращения молекулы не произойдет, а если он слишком велик, то превращение осуществляется, но расход энергии будет непродуктивным. В процессе фотосинтеза имеется ряд промежуточных реакций, которые происходят до момента образования углевода. На каждую из этих реакций также расходуется определенное количество энергии. Продуктивность фотохимического процесса определяется количеством: квантов, а не величиной отдельного кванта.
Таблица 8. Характеристика отдельных участков спектра
|
|
|
|
Энергия |
Свет |
Типичная |
|
эВ на |
кДж на |
|
длина |
Частота, Гц |
1 моль |
|
|
1 квант |
|||
|
волны, мм |
|
квантов |
|
|
|
|
||
Ультрафиолетовый |
259 |
11,8×1014 |
4,88 |
470 |
Фиолетовый |
410 |
7,31×1О14 |
3,02 |
290 |
Синий |
460 |
6,52×1О14 |
2,70 |
260 |
Зеленый |
520 |
5,77×1О14 |
2,39 |
230 |
Желтый |
580 |
5,17×1О14 |
2,14 |
206 |
Оранжевый |
620 |
4,94×1О14 |
2,00 |
193 |
Красный |
680 |
4,41×1О14 |
1,82 |
176 |
Инфракрасный |
1400 |
2,14×1О14 |
0,88 |
85 |
Энергия света Е, переносимая фотоном, или квантом hv, равна:

где h — световая константа Планка, равная 6,26196×10-34 Дж/с; v — частота колебаний данной световой волны, Гц; λ — длина волны, нм; с — скорость света в вакууме (примерно 3×108 м/с).
Согласно приведенному уравнению, квант света обладает энергией, пропорциональной его частоте и обратно пропорциональной длине волны.
В фотохимии употребляется единица количества света — эйнштейн . Она равна 6,023×1023 квантов монохроматического света. При поглощении системой, способной к фотохимическим
превращениям, |
происходит фотохимическое превращение 6,023×1023 молекул, или одного |
|
моля вещества. Энергия |
равна 6,023×1023 hv квантов, или фотонов, и называют ее молем |
|
квантов. |
|
|
В таблице 8 приведена характеристика участков спектра от ультрафиолетовых (видимых) до инфракрасных лучей.
Как установлено О. Варбургом и другими учеными, квантовый выход фотосинтеза (φ), т. е. количество молекул реагирующего углекислого газа на квант света, который поглощается при фотосинтезе, равен приблизительно 0,25. Следовательно, на восстановление одной молекулы СО2 расходуется 4 кванта красного света:
Количество молекул воспринятого |
или выделенного |
Число поглощенных квантов
— квантовый выход < 1.
Однако многочисленные экспериментальные определения квантовых выходов фотосинтеза показали, что при восстановлении одной молекулы СО2 поглощается 8-12 квантов света.
Поглощение 4 квантов красных лучей может дать достаточное и даже избыточное количество энергии, а именно: 4 кванта красного света дают 502-669 кДж/моль, а для восстановления 1 моля СО2 необходимо 470 кДж.
Очевидно, из 12 квантов энергия 4 квантов превращается в химическую энергию и связывается в продуктах фотосинтеза, энергия остальных 8 квантов используется на образование каких-то неустойчивых, высокоэнергетических продуктов, которые в дальнейшем, подвергаясь экзэргоническим превращениям, отдают энергию на поддержание необходимых для фотосинтеза градиентов потенциалов; потом эта энергия деградирует в тепло.
Обратная квантовому выходу величина — квантовые расходы |
|
должно быть поглощено |
|
растением для поглощения и превращения одной молекулы СО2 или выделения молекулы О2.
При очень низкой интенсивности света расход на фотосинтез не превышает 4 квантов. При увеличении интенсивности света количество квантов на восстановление одной молекулы СО2
резко возрастает. При 12 квантах коэффициент полезного действия энергии активного света, который поглощается фотосинтетически, может составлять 19%, или 8-10% энергии общей радиации солнца, падающей на лист. Считают, что листья зеленых растений обычно поглощают 80-85% энергии видимой части солнечного спектра, пропускают 5-10 и отражают около 10%.
Фотосинтез является окислительно-восстановительным процессом, в котором вода служит восстановителем и сама окисляется, а углекислый газ — окислителем и сам восстанавливается. В процессе фотосинтеза хлорофилл играет роль фотосенсибилизатора, т. е. вещества, поглощающего свет, с помощью энергии которого осуществляются химические превращения других веществ.
При поглощении квантов света хлорофилл приобретает качество активного окислителя и становится способным окислять некоторые вещества, отнимая у них электрон или водород — происходит реакция фотовосстановления хлорофилла с запасанием энергии квантов света в продуктах реакции. Такой хлорофилл способен активно восстанавливать ряд окислителей. Реакцию обратимого фотовосстановления хлорофилла, которая получила в литературе название реакции Красновского, можно представить следующими схемами (1, 2, 3):
хлорофилл квант возбужденный хлорофилл света в бирадикальном состоянии
Возбужденная молекула хлорофилла воспринимает электрон от молекулы донора с образованием пары первичных ион-радикалов:
Акт восприятия электрона является первичным фотохимическим процессом. Далее происходит реакция переноса протона (водорода) на молекулу хлорофилла:
Важная особенность процесса фотосинтеза заключается в том, что под влиянием энергии света происходит окисление воды и восстановление, ее водородом углекислого газа. Химической основой этого процесса является перенос электрона и протона от воды к СО2. Четыре молекулы
хлорофилла |
поглощают 4 кванта лучистой энергии. |
Хлорофилл, поглотивший 4 кванта, — возбужденный хлорофилл (обозначен звездочкой): под влиянием лучистой энергии, поглощенной хлорофиллом, происходит
фотолиз воды:
Опыты А. П. Виноградова и Р. В. Тейс показали, что изотопный состав кислорода, выделяющегося при фотосинтезе, соответствует соотношению 16О, 17О, 18О, что характерно для «обычной» воды. Опытами С. Рубена и М. Камена с разным содержанием, 18О в Н2О и СО2, которые давали
водоросли хлорелле на свету, установлено, что изотопный состав углекислого газа не влиял на изотопный состав кислорода, выделявшегося хлореллой, но соответствовал изотопному составу молекул воды, которую усваивала водоросль. Таким образом, кислород, выделяющийся при фотосинтезе, принадлежит воде.
В реакциях фотосинтеза углекислый газ выступает в виде карбоксильной группы, характерной для органических кислот. Затем, осуществляется реакция фотолиза — разложения воды под влиянием лучистой энергии, поглощаемой хлорофиллом. Наконец, за счет водорода,

освобождающегося при фотолизе воды, происходит восстановление углекислого газа до углевода.
Реакция превращения световой энергии в химическую связывается с реакциями восстановления хлорофилла, окислением воды в реакции Хилла и фосфорилированием с образованием АТФ. Изучение фотохимической реакции у изолированных хлоропластов начал Р. Хилл в 1937 г., когда было обнаружено, что суспензия хлоропластов на свету без участия СО2 способна выделять
кислород (А — акцептор водорода):
хлоропласт
свет
В основе этой реакции лежит фотолиз воды при непосредственном участии молекул хлорофилла. На основании реакции Хилла разработан метод определения фотохимической активности хлоропластов. Суть его заключается в том, что к суспензии хлоропластов или гомогенату листьев добавляют краску 2,6-дихлорфенолиндофенол или другой акцептор электрона, например феррицианид калия. По изменению окраски суспензии хлоропластов на свету (сравнивают с темновым вариантам) судят о фотохимической активности хлоропластов.
Исследованиями, проведенными на кафедре физиологии и биохимии растений Украинской сельскохозяйственной академии Л. Г. Литвиненко с изолированными хлоропластами из растений озимой пшеницы, кукурузы, подсолнечника и гороха, установлена связь между фотохимической активностью хлоропластов и первичными фотопроцессами в них — флуоресценцией и образованием парамагнитных центров. Как показали экспериментальные данные, реакция Хилла является чувствительным фотосинтетическим параметром, который можно использовать для характеристики световой фазы процесса фотосинтеза.
Таким образом, основные этапы фотосинтеза следующие:
1.Поглощение пигментами листа фотонов, несущих световую энергию, перенос ее на АДФ, которая превращается в АТФ, — происходит фотосинтетическое фосфорилирование.
2. Фотолиз воды (реакция Хилла): |
перенос водорода на НАДФ, который |
восстанавливается до НАДФН2, и освобождение кислорода.
3.Поглощение и перенос СО2 на сахар рибулозо-1,5-дифосфат с образованием двух молекул фосфоглицериновой кислоты (ФГК).
4.Присоединение к ФГК водорода и восстановление ее до фосфоглицеринового альдегида, который полимеризуется до гексозы, при этом регенерируется рибулоза.
Исследования последних лет (Д. Арнон и др.) показали, что первичными стабильными продуктами фотосинтеза, в которых аккумулируется энергия поглощенного кванта света и водород, являются АТФ и восстановленный никотинамидадениидинуклеотидфосфат (НАДФН2).
Они образуются в процессе двух реакций циклического (1) и нециклического (2) фотосинтетического фосфорилирования и представляют собой компоненты «восстановительной силы», которая обусловливает восстановление СО2 и образование углеводов (3) в растении. Эти
реакции можно схематически записать в виде уравнений:
АДФ |
|
|
АТФ |
|
НАДФ |
АДФ |
|
|
|
НАДФН |
АТФ |
|||
|
НАДФН |
АТФ |
|
|
|
|
|||
НАДФ АДФ Ф фосфор
Доказано, что количество АТФ, образовавшейся при фотофосфорилировании, соответствует количеству восстановленного окислителя в реакции Хилла и что этот процесс связан с пигментами — хлорофиллами и каротиноидами. Возможно, что ксантофиллы, в частности, участвуют в переносе кислорода при фотоокислении воды в реакциях фосфорилирования.
Американские ученые Эмерсон и Льюис установили, что квантовый выход фотосинтеза хлореллы (т. е. количество молекул СО2, которые реагируют на квант поглощенного света при фотосинтезе)
начинает падать около 680 нм и достигает нуля около 700 нм, хотя полоса поглощения хлорофилла α кончается лишь около 820 нм. Однако низкий квантовый выход фотосинтеза в области «красного падения» можно увеличить и даже довести до нормального уровня одновременным освещением светом более короткой длины волны. Это явление назвали эффектом Эмерсона. Спектр действия данного эффекта параллелен кривой, которая показывает долю поглощения света, приходившуюся в хлорелле на хлорофилл b, у диатомовых водорослей на фукоксантин и хлорофилл с, а у красных и сине-зеленых водорослей — на фикобилины. Очевидно, фотосинтез требует одновременного возбуждения хлорофилла и одного из вспомогательных пигментов. Световая энергия, поглощенная пигментами-спутниками, передается резонансно на хлорофилл α, и эффективность этого переноса определяет также действенность света, поглощенного вспомогательным пигментом, сенсибилизирующим фотосинтез. Таким образом, для эффективного использования световой энергии в фотосинтезе, кроме хлорофилла α, должен активироваться еще и вспомогательный пигмент — хлорофилл b, а также фикобилины, каротиноиды.
Общепризнано, что световая энергия, необходимая для реакций фотосинтеза, поступает на двух участках фотосинтетической электронно-транспортной цепи (ЭТЦ) с помощью двух последовательных фотохимических реакций, с участием поглощающих свет пигментов. Принципиальная сущность фотосинтеза заключается в отторжении электрона от молекулы воды и переносе его с помощью световой энергии, поглощенной пигментами, на СО2, который и
восстанавливается до углеводов.
Согласно современным представлениям о процессе фотосинтеза, большинство молекул пигментов фотосинтетического аппарата выполняют лишь обслуживающую роль акцепторов квантов света, передающих поглощенную энергию на реакционные центры, в которых происходят фотохимические реакции, связанные с восстановлением углекислого газа (рис. 30).
В основе механизма фотосинтеза лежат две фотохимические реакции, в которых создается «восстановительная сила» (АТФ и НАДФН2) с выделением кислорода, с участием не менее двух
форм хлорофилла α и b и, вероятно, других пигментов хлоропластов, поглощающих свет различной длины волны.
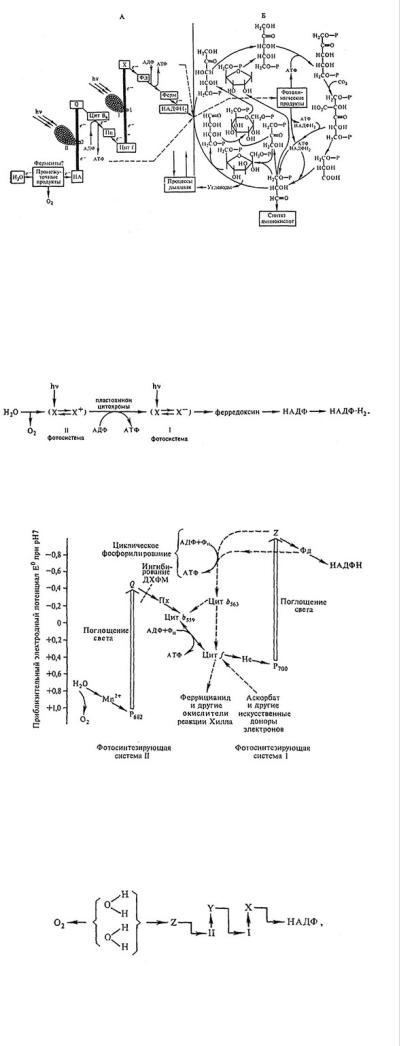
Рис. 30. Схема процессов фотосинтеза в хлоропласте (по В. Б. Евстигнееву): A — световая фаза — транспорт электрона; Б — темновая фаза — цикл ассимиляции углерода (по М. Кальвину); I, II — первая и вторая фотохимические системы фотосинтеза, включающие комплекс фотосинтетических пигментов; 1, 2 —
соответствующие им реакционные центры; е- — электрон; НА — донор электрона для фотосистемы II; Q — пластохинон; Цит — цитохром; Пц — пластоцианин*; X — неидентифицированный акцептор электрона от фотосистемы I; Фд — ферредоксин; ферм — ферредоксин-НАДФ-редуктаза; НАДФН2 — восстановленный никотинамидадениндинуклеотидфосфат; АДФ — аденозиндифосфат; АТФ — аденозинтрифосфат. Вертикальными жирными линиями обозначены процессы, идущие с накоплением световой энергии.
* Пластоцианин — это медьсодержащий белок, состоящий из одной полипептидной цепочки с молекулярной массой около 10 500. Одна молекула пластоцианина приходится на 400 молекул хлорофилла.
Управляющие функции хлорофиллового аппарата при фотосинтезе, переход энергии кванта в форму химической связи, наличие двух пигментных систем, двух фотохимических реакций фотосинтеза и образование основных компонентов «восстановительной силы» при нециклическом фотофосфорилировании могут быть записаны следующим образом:
Реакции, связанные с транспортом и превращением световой энергии в химическую, происходят в тилакоидах гран и межгранальных тилакоидах. Фосфорилирование при циклическом транспорте электронов сопряжено с фотосистемой I, а не циклический электронный транспорт происходит с последующим участием I и II фотосистем. Циклическая и нециклическая системы электронного транспорта осуществляются последовательно или параллельно.
Рис. 31. Z-схема (от слова zigzag) фотосинтеза (фотовосстановление в хлоропластах): P682 и Р700
— хлорофиллы реакционных центров; Q — предполагаемый первый акцептор электронов; Пх — пластохинон; Пс — пластоцианин; Z — первый акцептор электронов в фотосистеме I: Цит b, цит f — цитохромы b, f, Фд — ферредоксин; ДХФМ — дихлорфенилдиметилмочевина, блокирующая перенос электронов между двумя фотосистемами; Фн — фосфор неорганический. Штриховой стрелкой показан путь электронов, находящихся на
«вершине» фотосистемы I, возвращающихся в замыкаемый цикл. Е° — нормальный окислительновосстановительный потенциал при pH 7.
В настоящее время широко принята двухквантовая Z-схема (от слова zigzag) фотосинтеза (рис. 31). При этой схеме для переноса одного электрона или в конечном результате атома Н от воды к НАДФ необходимо два кванта света:
где 1Н — 2 кванта |
X, Y — акцепторы электрона пигментной системы I и II; Z — |
|
вещество, способное окислять воду. |
Таким образом, световой период процесса фотосинтеза представляется следующим образом. Энергия света, поглощенная молекулами хлорофилла, используется на разложение воды, в результате чего выделяется газообразный кислород и освобождаются электроны (водород), которые через сложную цепь промежуточных переносчиков (передатчиков) закрепляются в НАДФН2. В цепи переноса электронов участвуют пластохинон и ферредоксин. Пластохинон
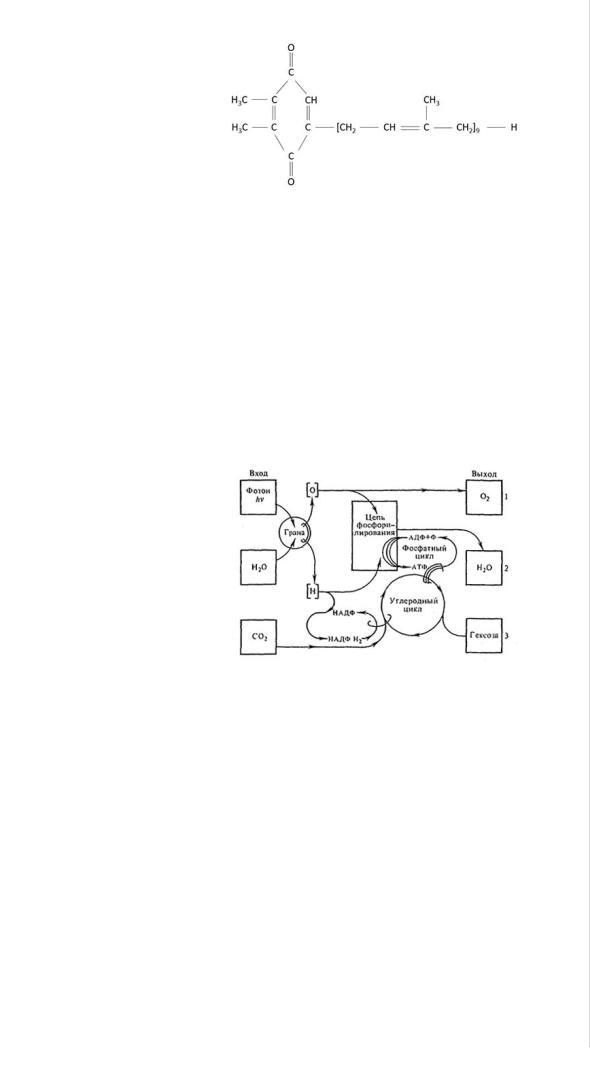
является производным бензохинона (2,3-диметил-5-соланезилбензохинон) и имеет следующее строение:
Ферредоксин (ФД) — один из восстановленных кофакторов, который также локализован в хлоропластах; это белок с небольшой молекулярной массой (12000), содержащий железо в негеминовой форме. Ферредоксины обнаружены у бактерий, водорослей и высших растений. Для некоторых из них установлена первичная структура.
Ферредоксин имеет низкий окислительно-восстановительный потенциал (при pH 7-0,42 В), что позволяет ему быть переносчиком электронов: он принимает электроны из фотосистемы I. Восстановленный ферредоксин хлоропластов вместе со специфическим ферментом вызывает восстановление НАДФ до НАДФН2 (рис. 32).
Согласно приведенным схемам (см. рис. 30, 32), где объединены световая и темновая фазы процесса фотосинтеза в хлоропласте, молекулы фотосистемы II, поглощая квант света, переводят электрон в возбужденное состояние, который и воспринимается пластохиноном. Молекулы хлорофилла реакционного центра фотосистемы II вместо утраченного электрона присоединяют электрон молекулы воды или радикала ОН. Пластохинон при участии цитохромов и пластоцианина передает электрон реакционному центру фотосистемы I. Эта фотосистема, также поглощая квант света, отдает возбужденный электрон через катализатор ферредоксин и фермент — ферредоксин-НАДФ-редуктаза НАДФ, который акцептирует ион водорода, образуя НАДФН2 — восстановитель с высоким потенциалом.
Таким образом, вместо утраченного электрона окисленные молекулы хлорофилла фотосистемы I становятся активным окислителем и легко воспринимают электрон от молекул хлорофилла фотосистемы II через соответствующие каталитические системы. За счет энергии возбужденных электронов по пути их миграции из АДФ образуется АТФ. Кислород освобождается при фотолизе воды, химизм которого мало изучен.
Рис. 32. Блок-схема реакций в хлоропласте: 1 — на одну поглощенную молекулу СО2 освобождается одна молекула О2; 2 — на одну ассимилированную молекулу СО2 приходится одна молекула Н20 (суммарный расход): 3 — суммарный выход: одна гексоза на шесть использованных молекул СО2.
Итак, образование «восстановительной силы» АТФ и НАДФН2 связано с миграцией электрона молекул хлорофилла в I и II пигментно-ферментных фотохимических системах хлоропласта.
У высших растений имеется два типа реакционных центров. Один из них представлен более длинноволновыми формами хлорофилла α 683 и 700 нм (фотосистема I). В фотосистеме II роль реакционного центра выполняют более коротковолновая форма хлорофилла α — 670 нм, некоторые каротиноиды и фикобилины. Всего в одном хлоропласте может содержаться в
среднем около 2×106 реакционных центров. Исследования локализации фотосистем в мембранах хлоропластов и разделение их на фракции с применением метода ферментативного гидролиза и дифференциального центрифугирования показали, что более тяжелые частицы (при 20000 g*) оказались обогащенными компонентами II фотосистемы, а более легкие (выделенные при 145000 g)— I фотосистемы.
*Гравитационная единица, показывающая, во сколько раз центробежная сила в центрифуге больше силы земного притяжения.
Исследования тонкой структуры фотосинтетических мембран хлоропластов гороха и кукурузы (Л. К. Островская и др.) показали, что фракция 145000 g содержит в основном частицы диаметром около 10 нм и имеет вид электронно-прозрачных колец с электронно-плотными ядрами внутри или без них, кольца состоят из пяти субъединиц размером около 3 нм каждая. Фракция 20000 g представляет собой диски, состоящие также из субъединиц. Установлено, что мембраны межгранальных тилакоидов содержат частицы, соответствующие фотосистеме. I (легкие), тогда как мембраны гран имеют частицы, характерные для I и II фотосистем.
Исследования спектральных свойств фрагментов мембран, фосфорилирующей и АТФ-азной активности позволили предположить, что I и II фотосистемы и соответствующие фотохимические реакции определенным образом пространственно локализованы (вмонтированы) на мембранах, но в то же время они тесно сопряжены между собой, поэтому указанные фрагменты мембран в литературе получили наименование сопрягающий фактор I и сопрягающий фактор II фотосистемы.
Фотосинтез, как и все биологические окислительно-восстановительные реакции, сопряженные с накоплением энергии, относятся к двухэлектронным системам. Существованием таких систем

объясняется восстановление НАДФ до НАДФНд и образование АТФ — «восстановительной силы» в терминальной реакции световой фазы фотосинтеза.
Фотосинтетическая единица (ФЕ) — это комплекс пигментов и других молекул, состоящий из светособирающих пигментов — антенны и реакционного центра, осуществляющего перенос электрона. У высших растений фотосинтетическая единица включает 250-300 молекул хлорофилла и 50 молекул каротиноидов. Реакционный центр состоит из белка (трех субъединиц), двух молекул хлорофилла Р730 И Р700. Двух молекул феофитина, цитохромов, ферредоксина и
НАД.
Передача энергии в фотосинтетической единице осуществляется путем миграции ее от молекулы к молекуле — так называемой резонансной миграции энергии при слабых диполь-дипольных взаимодействиях молекул. Передача энергии такого вида возможна только при расстоянии между молекулами, не превышающем 10 нм. Резонансная миграция энергии между молекулами пигментов разных видов называется гетерогенной (например, хлорофилл — каротиноиды), а перенос энергии между тождественными молекулами хлорофилла α — гомогенной.
Считается, что поглощение квантов света происходит последовательно каждой из молекул пигмента — на прямом солнечном свету всего один раз за 0,1 с. Затем энергия поглощенных фотонов стекается к реакционному центру, где она превращается в химическую энергию органических соединений: «Пропускная» способность фотосинтетической единицы (насыщение фотосинтеза) соответствует таким степеням освещенности, при которых реакционный центр поглощает (перерабатывает) 50 квантов в 1 с.
Энергия квантов солнечного излучения, поглощаемая основной массой хлорофилла — светособирающими молекулами хлорофилла (антенна), мигрирует к реакционным центрам, которые непосредственно участвуют в фотосинтетическом переносе электрона:
Исследования показали, что в первой фотосистеме ста молекулам светособирающего хлорофилла соответствует одна молекула хлорофилла реакционного центра. Реакционные центры, выделенные из хроматофоров фотосинтезирующих бактерий, включают белок (3 субъединицы), бактериохлорофилл (4 молекулы), бактериофеофитин (2 молекулы), первичные акцепторы уби- и пластохинон (1-2 молекулы) и цитохромы — доноры электрона. Пигменты в реакционных центрах находятся преимущественно в агрегированных формах. Установлено, что пигменты реакционных центров глубже погружены внутрь мембраны по сравнению с более поверхностным расположением светособирающих молекул хлорофилла. У фотосинтезирующих бактерий в реакционных центрах донорами электронов являются бактериохлорофилл 960 нм и 890 нм и акцепторами — убихинон и Fe.
У зеленых растений для I фотосистемы донорами электронов служат цитохромы и пигменты Р*700 и Р*730, для II фотосистемы — вода и Р*680; акцепторами электронов для обеих фотосистем
являются пластохиноны.
*Хлорофиллы реакционных центров
Исследования показали, что возбужденная молекула хлорофилла обладает склонностью к отдаче электрона, а возбужденная молекула феофитина, безмагниевый аналог хлорофилла, — к его восприятию. Такой обратимый перенос электрона лежит в основе действия пигментов в реакционных центрах (по А. А. Красиовскому):
Установлено, что Мn способствует восстановлению феофитина. Следовательно, переход энергии фотона в форму химических связей обусловлен работой управляющего устройства фотосинтетического аппарата. Управляющие функции хлорофилла при фотосинтезе, его фотохимические реакции служат источником энергии для образования органических соединений.
Плотная упаковка хлорофилла на фотосинтетических мембранах дает основание рассматривать фотосинтез как фотофизический процесс. Квант света, поглощенный молекулой хлорофилла, передает свою энергию электрону, который переходит в возбужденное состояние. Если при этом спин* электрона не изменяется, то он будет недолговременным — это так называемое синглетное состояние. Если же спин электрона изменяется, то возникает метастабильное триплетное
состояние электрона. Продолжительность такого состояния значительно больше (10-2 с), чем
синглетного (10-9 с), поэтому триплетное состояние электрона играет основную роль в процессе фотосинтеза (рис. 33).
*Спин - собственный механический момент количества движения электрона.
Возбужденный электрон может перейти в основное состояние с излучением кванта света, т. е. сопровождается флуоресценцией. Молекула хлорофилла, находясь в триплетном возбужденном состоянии, имеет два валентных электрона с неспаренными спинами и представляет собой бирадикал. Возбужденный электрон принадлежит уже не отдельной молекуле, а системе молекул, которые могут переходить из одной зоны уровней в другую. На том месте, откуда электрон перешел в возбужденное состояние, остается вакансия; она обладает свойством положительного заряда и называется «дырка». Возбужденный электрон и «дырка» образуют пару,
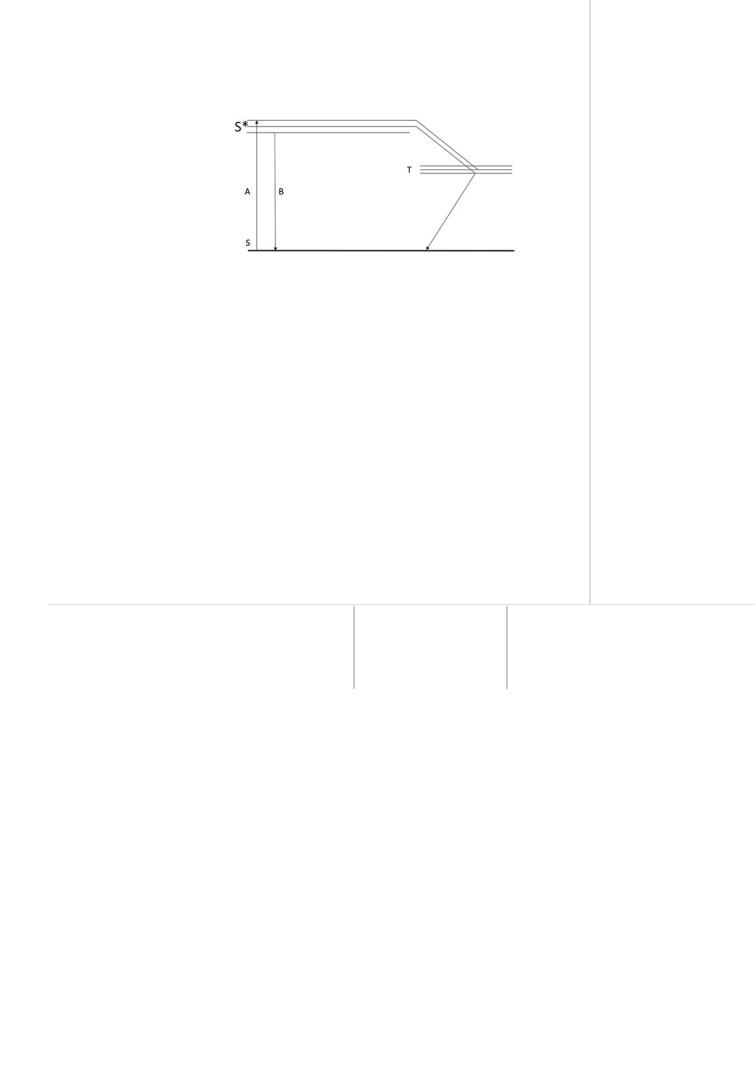
которую называют экситоном — он может мигрировать по зонам уровней, перенося с собой энергию поглощенного кванта света. Места в зонах, где «застревает» мигрирующий электрон, называют ловушками. Доноры электронов могут отдавать их «дыркам», а акцепторы электронов могут поглощать их из ловушек. В результате переноса энергии электронного возбуждения образуются вещества с высокоэнергетическими связями — АТФ и НАДФН2, энергия которых
используется для восстановления СО2 в процессе фотосинтеза.
Протонный цикл в хлоропластах. Хлоропласты — это биологические аккумуляторы, зарядка АТФ которых может происходить по принципу хемоосмотического механизма, предложенного П. Митчеллом для окислительного фосфорилирования.
При действии света на хлоропласт начинается электронный поток по системе переносчиков, встроенных в ламеллы тилакоидов. С переносом электронов по ЭТЦ сопряжено поступление протонов из окружающей среды внутрь тилакоидов. При этом, кроме переносчиков электронов, должны присутствовать доноры и акцепторы протонов. В таком случае компоненты среды могут
присоединять Н+ на одной стороне мембраны и при окислении отдавать на другую сторону. В результате в хлоропласте создаются мембранный протонный градиент и разность потенциалов на мембранах (достигающая 50 мВ), обеспечивающая при участии обратимой АТФ-азы образование АТФ.
Существование такого механизма подтверждается опытами с изолированными хлоропластами, которые в полной темноте могут образовывать АТФ. Для этого хлоропласты сначала инкубировали в среде с pH 4, а затем помещали в среду с pH 8,5, содержащую АДФ, неорганический фосфор и ингибитор возможного потока электронов. Образование АТФ И. А. Тарчевский объясняет следующим образом. При pH 4 протоны медленно проникали через мембрану внутрь хлоропласта до выравнивания pH внутри и вне хлоропластов. Затем быстрое перенесение хлоропластов в среду с pH 8,5 создавало искусственный трансмембранный протонный градиент, достаточный для образования АТФ.
Таким образом, протонодвижущая сила, или разность электрохимических потенциалов водородных ионов , обусловила образование макроэргической связи в молекуле АТФ,
Группой исследователей Франции, США и Великобритании под руководством Т. Мура (Аризонский университет, США) в 1984 г. синтезирован молекулярный комплекс, который с большой точностью имитирует основные процессы природного фотосинтеза.
В состав молекулярного комплекса входят: порфирин (по своему составу родствен хлорофиллу), поглощающий свет; хинон, захватывающий возбужденные в порфирине электроны; β-каротин который пополняет электронами «дырки», образующиеся в порфирине в результате захвата хиноном возбужденных электронов, а также расширяет область длин воли в коротковолновой части спектра, поглощаемых порфирином. В молекулярном комплексе отрицательный заряд концентрируется в хиноне, а положительный собирается на β-каротине. Разделенные заряды
обладают относительно долгим сроком жизни — 3×10-6 с по сравнению с 3×10-12 с для системы порфирин — хинон.

Фотосинтез