

Osnovy_zoologii
.pdf681
Рисунок 34.2 – Бактерии: стрептококки и бациллы
Автотрофность
Следующая ступень (ароморфоз) развития жизни на Земле – появление фотосинтетических пигментов в клетках прокариот. Эти прокариоты – цианобактерии (сине-зеленые водоросли, цианеи) (рисунок 34.3) – сами синтезировали органические вещества из неорганических
(процесс фотосинтеза), т.е. были автотрофами.
Рисунок 34.3 – Цианобактерии: колония ностока под микроскопом и цианобактерии
в аквариуме
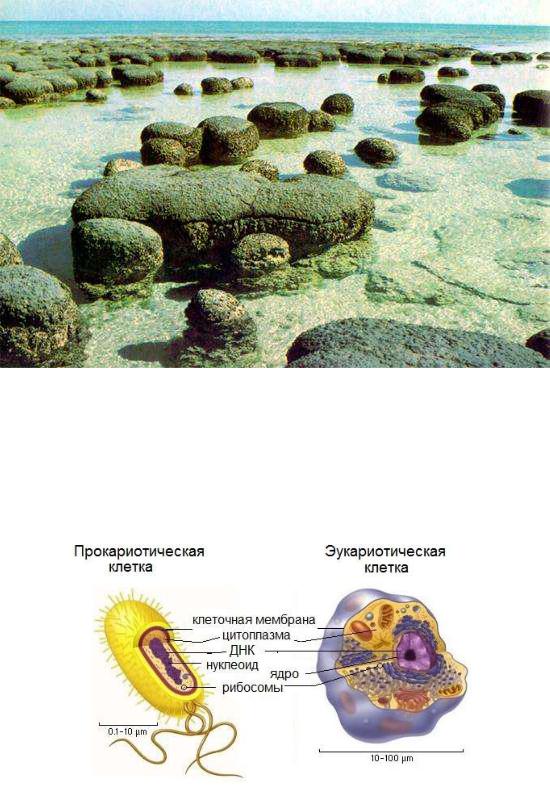
Самые ранние свидетельства жизни на Земле – окаменевшие продукты жизнедеятельности сине-зеленых водорослей – строматолиты (рисунок 34.4). Самые ранние относятся к архею (примерно 3, 5 млрд. лет назад).
682
Рисунок 34.4 – Строматолиты
Ядерность
Следующий ароморфоз произошел на границе Архея и Протерозоя (2,6
– 2,7 млрд. лет назад): появление в клетках ядра. Организмы, в клетках которых есть ядро, называются эукариотами (рисунок 34.5).
Рисунок 34.5 – Сравнение прокариотической (бактериальной) и эукариотической
клетки
После формирования клеточного ядра все эукариотические организмы разделились на две ветви: эукариотических автотрофов (предки растений) и эукариотических гетеротрофов (предки грибов и животных).
683
Колониальность
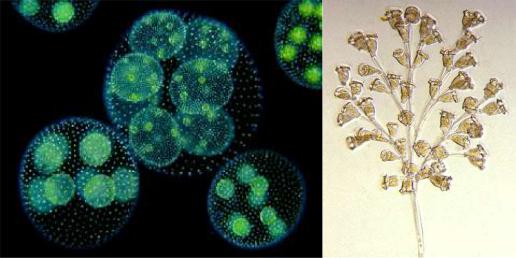
Следующий шаг: появление колониальных одноклеточных эукариот (рисунок 34.6).
Рисунок 34.6 – Колониальный автотроф – вольвокс; колониальный гетеротроф –
инфузория кархезиум
Биологический смысл колониальности:
более эффективная защита от врагов;
защита внутренних клеток колонии от неблагоприятных факторов внешней среды:
более активная добыча питательных веществ.
Многоклеточность
Следующим ароморфозом было происхождение многоклеточности. Этот ароморфоз произошел в конце Протерозоя.
Значение многоклеточности:
специализация клеток и более эффективное выполнение функций;
увеличение размеров: выгодно при защите от хищников и при добыче пищи;
более быстрое передвижение;
увеличение сроков жизни благодаря регенерации.
Существует несколько теорий происхождения многоклеточности: теория фагоцителлы Мечникова, теория гастреи Геккеля и др. (рисунок 34.7).

684
Рисунок 34.7 – Происхождение многоклеточности
Эволюция многоклеточных животных
Первыми многоклеточными животными были радиальносимметричные организмы: губки и кишечнополостные (рисунок 34.8). Все они вели прикрепленный образ жизни.
Рисунок 34.8 – Радиальная симметрия и радиально-симметричные животные
Далее появились двусторонне-симметричные активно передвигающиеся животные: плоские (рисунок 34.9) и круглые черви. Они имели головной конец тела, на котором концентрировались органы чувств.

685
Рисунок 34.9 – Двусторонняя (билатеральная) симметрия и плоский червь планария
Вендский период
Примерно 650 млн. лет назад Землю населяли мягкотелые существа – вендобионты – первые известные многоклеточные животные: губки, медузы, плоские черви (рисунок 34.10). Они были мягкотелыми, поэтому остатки их плохо сохранились.
Рисунок 34.10 – Вендский период (630 млн. лет назад)
Кембрийский период
В кембрии (540 млн. лет назад) появились хищные организмы и средства нападения и защиты: челюсти, панцири, раковины. Многие животные имели твердый наружный скелет, поэтому сохранилось большое количество остатков кембрийской фауны (рисунок 34.11). В связи с этим ученые назвали этот период «кембрийским взрывом».

686
Рисунок 34.11 – Кембрийский период (540 млн. лет назад)
От древних ресничных червей произошли кольчатые черви. Древние морские многощетинковые кольчатые черви, вероятно, послужили основой для возникновения типов членистоногих, моллюсков и хордовых (рисунок
34.12, 34.13, 34.14, 34.15).
Расцвет трилобитов – ближайших предков ракообразных.
Появление гигантских хищных ракообразных – аномалокарид (лаггания, аномалокарис, хардия)
Рисунок 34.12 – Остатки трилобита и реконструкция лаггании

687
Рисунок 34.13 – Так ученые реконструировали хардию – нового представителя
аномалокарид, удивительных хищников кембрия
Рисунок 34.14 – Кембрийские билатерально-симметричные животные
Рисунок 34.15 – Кембрийская Виваксия – возможный предок кольчатых червей

688
Первые рыбоподобные позвоночные животные – остракодермы – появились в конце кембрия. Они были покрыты панцирем из костных щитков и не имели челюстей. До наших дней дожили только паразитические представители бесчелюстных – миноги и миксины.
Силур
Период известен массовым вымиранием, в результате которого исчезло около 60 % видов существовавших в ордовике морских организмов.
Появлись акантоды (колючкозубые рыбы) (рисунок 34.16) и челюстноротые рыбы – костнопанцирные и беспанцирные.
Рисунок 34.16 – Акантода климатиус
Увеличивается видовое разнообразие прямораковинных головоногих моллюсков и граптолитов (рисунок 34.17, 34.18). Брахиоподы (плеченогие) увеличили свое разнообразие в три раза, в силуре к ним относится 8% всех родов.
Рисунок 34.17 – Гигантский камероцерас и ракоскорпион

689
Рисунок 34.18 – Планктонный граптолит Loganograptus и брахиопода
Megamastax amblyodus из позднего силура, костная рыба длиной до метра, на 2014 год считается первым позвоночным хищником, специализирующимся на поедании других позвоночных (рисунок 34.19).
Рисунок 34.19 – Megamastax amblyodus и его предполагаемые жертвы –
панцирные бесчелюстные Dunyu longiforus
Девон
Процветают бесчелюстные панцирные остракодермы (цефаласписы и др.) (рисунок 34.20).
Появились и быстро завоевали водные пространства головоногие моллюски аммониты, позже вымершие вместе с динозаврами в меловой период.

690
Рисунок 34.20 – Цефаласпис и аммонит
В девоне начался расцвет челюстноротых. Для большинства этих животных характерны наличие двусторонней симметрии, третьего зародышевого листка (мезодермы), полости тела, наружного (членистоногие) или внутреннего (хордовые) твердого скелета, прогрессирующая способность к активному передвижению, обособление переднего конца тела с ротовым отверстием и органами чувств, постепенное совершенствование центральной нервной системы.
От первых челюстноротых возникли лучеперые и кистеперые рыбы (рисунок 34.21). Кистеперые имели в плавниках опорные элементы, из которых позже развились конечности наземных позвоночных. Из жаберных дуг образовались подвижные челюсти, а из кожных складок – плавники. Формирование поясов парных грудных и брюшных конечностей способствовало увеличению маневренности движений.
Двоякодышащие и кистеперые рыбы посредством плавательных пузырей, имеющих связь с пищеводом и снабженных системой кровеносных сосудов, могли дышать атмосферным кислородом.