

Osnovy_zoologii
.pdf561
Предком черепах принято считать пермского эунотозавра (Eunotosaurus) – небольшое ящерообразное животное с короткими и очень широкими ребрами, образующими подобие спинного щита. Брюшного щита у него не было. Имелись зубы. Мезозойские черепахи первоначально были сухопутными и, видимо, роющими животными. Лишь впоследствии некоторые группы перешли к водному образу жизни и в связи с этим многие из них частично утратили костный и роговой панцирь.
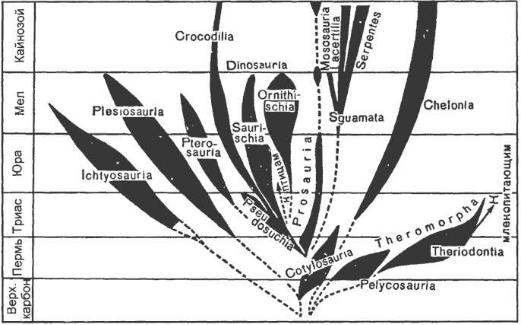
С триаса и до наших дней черепахи сохранили основные черты своей организации. Они пережили все испытания, которые погубили большинство рептилий, и в настоящее время так же процветают, как и в мезозое. Нынешние скрытошейные и бокошейные в большей мере хранят первичный облик сухопутных черепах триаса. Морские и мягкокожистые появились в позднем мезозое. Все прочие рептилии, как древние, так и современные, приобретали в структуре черепа одну или две височные впадины (рисунок 29.1). Одну, нижнюю, височную впадину имели синапсидные. Одна верхняя височная впадина отмечена в двух группах: параноидные и эвриансидные. И наконец, две впадины имели диапсидные.
Эволюционная судьба этих групп различна. Первыми от предкового ствола отошли синапсиды (Synapsida) – рептилии с нижними височными впадинами, ограниченными скуловой, чешуйчатой и заглазничной костями. Уже в позднем карбоне эта группа из первых амниот стала самой многочисленной. В палеонтологической летописи они представлены двумя последовательно существовавшими отрядами: пеликозавров (Pelicosauria) и терапсидов (Therapsida). Их еще именуют звероподобными (Theromorpha).
Рисунок 29.1 – Строение черепа: A –анапсид, B –синапсид, C –диапсид
Период своего расцвета звероподобные пережили задолго до того, как
562
появились первые динозавры, котилозавры были их непосредственными родичами. В частности, пеликозавры (Pelicosauria) были еще очень близки к котилозаврам. Их останки нашли в Северной Америке и в Европе. Внешностью они походили на ящериц и имели небольшие размеры – 1–2 м, имели двояковогнутые позвонки и хорошо сохранившиеся брюшные ребра. Однако зубы их сидели в альвеолах. У некоторых намечалась, хотя и в небольшой степени, дифференцировка зубов.
В средней перми пеликозавров сменили более высокоорганизованные зверозубые (Theriodontia): зубы у них были явственно дифференцированы, появилось вторичное костное нѐбо, единый затылочный мыщелок разделился на два, нижняя челюсть в основном была представлена зубной костью, положение конечностей также изменилось: локоть переместился назад, а колено – вперед, и в итоге конечности начали занимать положение под туловищем, а не по бокам его, как у других рептилий, в скелете появилось много черт, общих с млекопитающими.
Многочисленные пермские зверозубые рептилии были весьма разнообразны по облику и образу жизни. Многие были хищниками. Возможно, такой была иностранцевия (Inostranceviaalexandrovi). Другие питались растительной или смешанной пищей. Эти малоспециализированные виды стоят ближе всего к млекопитающим. Среди них надо указать на циногнатуса (Cynognathus), имевшего много прогрессивных черт организации.
Зверозубые были многочисленны еще в раннем триасе, но при появлении хищных динозавров они исчезли. Материалы, приведенные в таблице, свидетельствуют о резком сокращении разнообразия звероподобных на протяжении триаса. Звероподобные представляют большой интерес как группа, давшая начало млекопитающим.
Следующей группой, отделившейся от анапсидных котилозавров, были диапсидные (Diapsida). Их череп имеет две височные впадины, расположенные выше и ниже заглазничной кости. Диапсидные в конце палеозоя (пермь) дали чрезвычайно широкую адаптивную радиацию систематическим группам и видам, которых обнаруживают и среди вымерших форм, и среди нынешних рептилий. Среди диапсидных наметились две основные группы (инфра – классы): инфракласс Лепидозавроморфы (Lepidosauromorpha) и инфракласс Архозавроморфы (Archosauromorpha). Палеонтологи не располагают точными сведениями, для того чтобы сказать, кто из них древнее и моложе по времени появления, но эволюционная судьба их различна.
Лепидозавроморфы – этот древний инфракласс объединяет ныне живущих гаттерию, ящериц, змей, хамелеонов и их вымерших предков. Гаттерия, или сфенодон (Sphenodonpunctatus), живущая сейчас на небольших островах у побережья Новой Зеландии, – потомок достаточно распространенных в середине мезозоя первоящеров, или клинозубых (надотряд Prosauria, или Lepidontidae). Для них характерно множество
563
клиновидных зубов, сидящих на костях челюстей и на небе, как у амфибий, и амфицельные позвонки. Ящерицы, змеи и хамелеоны составляют ныне широкое многообразие отряда чешуйчатых (Squamata). Ящерицы – одни из древнейших продвинутых групп рептилий, их останки известны из верхней перми. Ученые обнаруживают много черт сходства ящериц и сфенодонов: их конечности широко расставлены, тело движется, волнообразно изгибая позвоночный столб, среди общих черт их морфологического сходства – присутствие интертарзального сустава.
Змеи появляются только в мелу. Хамелеоны – это специализированная группа более поздней эпохи – кайнозоя (палеоцен, миоцен).
Архозавров считают самыми удивительными из всех рептилий, когда- либо живших на Земле. Среди них – крокодилы, птерозавры, динозавры. Крокодилы – единственные архозавры, дожившие до наших дней. Появляются крокодилы в конце триаса. Юрские крокодилы существенно отличны от современных: отсутствием настоящего костного неба. Их внутренние ноздри открывались между небными костями. Позвонки были еще амфицельными.
Крокодилы современного типа с полностью развитым вторичным костным небом и процельными позвонками произошли от древних архозавров – псевдозухий. Известны они с мела (около 200 млн. лет назад).
Большинство обитало в пресных водоемах, но среди юрских форм известны и настоящие морские виды.
Крылатые ящеры, или птерозавры (Pterosauria), представляют собой один из замечательных примеров специализации мезозойских рептилий. Это были летающие животные весьма своеобразного строения: их крылья представляли складки кожи, натянутые между боками тела и очень длинным четвертым пальцем передних конечностей, широкая грудина имела хорошо развитый киль, как у птиц, кости черепа срастались рано; многие кости были пневматичными, вытянутые в клюв челюсти несли зубы, длина хвоста и форма крыльев варьировали: у одних (рамфоринхи) были длинные узкие крылья и длинный хвост, летали они, видимо, скользящим полетом, часто планируя; у других (птеродактили) хвост был очень короткий, а крылья широкие; полет их чаще был гребной.
Судя по тому, что остатки птерозавров обнаружены в отложениях соленых водоемов, это были обитатели побережий. Они питались рыбой и по поведению, видимо, были близки к чайкам и крачкам. Размеры варьировали от нескольких сантиметров до метра и более. Самые крупные среди летающих позвоночных принадлежат к позднемеловым крылатым ящерам. Это птеранодоны. Предполагаемый размах их крыльев 7-12 м, масса тела около 65 кг. Они обнаружены на всех континентах, кроме Антарктиды. Палеонтологи предполагают постепенное угасание в эволюции этой группы, что совпало по времени с появлением птиц.
Динозавры (Dinosauria) известны в палеонтологической летописи с середины триаса. Это наиболее многочисленная и разнообразная группа
564
рептилий, когда-либо живших на суше. Среди динозавров были мелкие животные, с длиной тела менее метра, и гиганты длиной почти до 30 м. Одни из них ходили только на задних лапах, другие – на всех четырех. Очень разнообразен был и общий облик, но у всех голова относительно тела была мала, а спинной мозг в крестцовой области образовывал местное расширение, объем которого превосходил объем головного мозга. В самом начале формирования динозавры разделились на две ветви, развитие которых шло параллельно. Характерной особенностью их было строение тазового пояса, в связи с чем эти группы именуются ящеротазовыми и птицетазовыми.
Ящеротазовые (Saurischia) первоначально были сравнительно мелкими хищными животными, передвигавшимися скачками только на задних ногах, в то время как передние служили для схватывания пищи. Для опоры служил и длинный хвост. В последующем появились крупные растительноядные формы, ходившие на всех четырех ногах. К ним относились самые крупные позвоночные, когда-либо жившие на суше: бронтозавр имел длину тела около 20 м, диплодок – до 26 м. Большинство гигантских ящеротазовых, видимо, были полуводными животными и питались сочной водной растительностью.
Птицетазовые (Ornithischia) получили свое название в связи с удлиненным тазом, похожим на таз птиц. Первоначально они передвигались на одних удлиненных задних ногах, но более поздние виды имели обе соразмерно развитые пары конечностей и ходили на четырех ногах. По характеру питания птицетазовые были исключительно растительноядными животными. Среди них – игуанодон, ходивший на задних ногах и достигавший в высоту 9 м. Трицератопс внешне был очень похож на носорога, обладал обычно небольшим рогом на конце морды и двумя длинными рогами над глазами. Длина его доходила до 8 м. Стегозавр отличался несоразмерно маленькой головой и двумя рядами высоких костных пластинок, располагавшихся на спине. Длина его тела была около 5 м.
Динозавры были распространены почти по всему земному шару и обитали в крайне разнообразных условиях. Они населяли пустыни, леса, болота. Некоторые вели полуводный образ жизни. Несомненно, что в мезозое эта группа рептилий была на суше господствующей. Наибольшего расцвета динозавры достигли в меловое время, а к концу этого периода вымерли.
Парапсидные и эвриапсидные – в черепе которых имелась только одна верхняя височная впадина. Высказано предположение, что они произошли от диапсидных путем утраты нижней впадины. В палеонтологической летописи они были представлены двумя группами: ихтиозавры (Ichthyosauria) и плезиозавры (Plesiosauria). В течение всего мезозоя, с раннего триаса до мела, они господствовали в морских биоценозах. Ихтиозавры (Ichthyosauria) занимали в мезозое то же место, которое ныне занимают китообразные. Они плавали, волнообразно изгибая тело, особенно хвостовую его часть, плавники у них служили для управления. Их конвергентное сходство с
565
дельфинами поразительно: веретенообразное тело, вытянутое рыло и большой двухлопастный плавник. Парные конечности у них превратились в ласты, при этом задние конечности и таз были недоразвиты. Фаланги пальцев были удлинены, а число пальцев у некоторых доходило до 8. Кожа была голой. Размеры тела варьировали от 1 до 14 м. Ихтиозавры жили только в воде и питались рыбой, отчасти беспозвоночными. Установлено, что они были живородящими. Ихтиозавры появились в триасе, в конце мела они вымерли.
Плезиозавры (Plesiosauria) имели иные, чем ихтиозавры, приспособительные особенности в связи с жизнью в море: широкое и плоское тело со сравнительно слаборазвитым хвостом. Орудием плавания служили мощные ласты. В отличие от ихтиозавров у них была хорошо развита шея, несущая небольшую голову. Их облик напоминал ластоногих. Размеры тела от 50 см до 15 м.Отличен был и образ жизни. Во всяком случае, некоторые виды населяли прибрежные воды. Питались рыбой и моллюсками.
Появившись в начале триаса, плезиозавры, как и ихтиозавры, вымерли в конце мелового периода.
Из приведенного краткого обзора филогении рептилий видно (рисунок 29.2), что подавляющее большинство крупных систематических групп (отрядов) вымерли до начала кайнозойской эры и современные рептилии представляют собой лишь жалкие остатки богатейшей мезозойской фауны рептилий. Причина этого грандиозного явления понятна лишь в самых общих чертах.
Рисунок 29.2 – Филогенетическое древо пресмыкающихся
Большинство мезозойских рептилий представляли собой чрезвычайно
566
специализированных животных. Успех их существования зависел от наличия весьма своеобразных жизненных условий. Надо думать, что односторонняя глубокая специализация была одной из предпосылок их исчезновения. Установлено, что хотя вымирание отдельных групп рептилий происходило в течение всего мезозоя, это проявлялось в конце мелового периода. В это время за сравнительно короткий срок вымерло большинство мезозойских пресмыкающихся. Если справедливо называть мезозой веком рептилий, то не менее оправданно название конца этой эры веком великого вымирания. Следует принять во внимание, что в меловое время произошли значительные изменения климата и ландшафтов. Это совпало с существенными перераспределениями суши и моря и движениями земной коры, приведшими к громадным горообразовательным явлениям, известным в геологии под названием альпийской стадии горообразования. Считают, что в это время вблизи Земли проходило крупное космическое тело. Нарушения сложившихся условий жизни в этой связи были весьма значительными. Однако они заключаются не только в изменении физического состояния Земли и других условий неживой природы.
В середине мелового периода произошла смена мезозойской флоры хвойных, саговниковых и других растений представителями флоры нового типа, а именно покрытосемянными. Не исключены и генетические изменения в природе самих рептилий. Естественно, что все это не могло не отразиться на успехе существования всех животных и специализированных в первую очередь.
Наконец, надо учесть, что к концу мезозоя все большее развитие получали несравненно более высокоорганизованные птицы и млекопитающие, сыгравшие важную роль в борьбе за существование между группами наземных животных.
567
ТЕМА 30: НАДКЛАСС ЧЕТВЕРОНОГИЕ, ИЛИ НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ– TETRAPODA. КЛАСС AVES – ПТИЦЫ
ПЛАН:
30.1Основные принципы организации систем органов птиц в связи с их адаптацией к полету
30.2Особенности высшей нервной деятельности, органы чувств и ориентация птиц в пространстве
30.3Размножение и характеристика эмбрионального и постэмбрионального развития
Прогрессивные черты организации птиц заключаются в более высоком уровне развития нервной системы, а в связи с этим и в более разнообразном и совершенном приспособительном поведении. В высокой и постоянной температуре тела, связанной со значительно возросшей интенсивностью обмена веществ и с более совершенной терморегуляцией .В способности к полету, что не ведет в подавляющем большинстве случаев к потере возможности передвигаться по твердому субстрату или лазать.В более совершенном размножении: насиживании яиц, обогреве, охране и выкармливании птенцов.
Птицы характеризуются большим, чем рептилии, многообразием и обилием видов. Класс включает в себя более 8 тыс. видов, объединенных в 35–40 отрядов.
30.1 Основные принципы организации систем органов птиц в связи с их адаптацией к полету
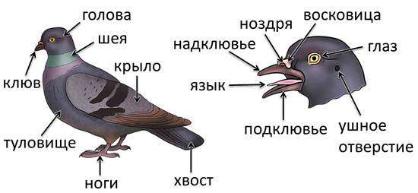
Внешнее строение птиц. Имеется голова, шея, туловище, конечности и хвост. Голова небольшая, округлая. Челюсти вытянуты в клюв и покрыты роговым чехлом. В клюве различают верхнюю часть – надклювье и нижнюю
– подклювье. У основания надклювья открываются ноздри. У голубя они прикрыты мягкой, белой, вздутой кожей – восковицей, которая выполняет осязательную функцию (рисунок 30.1). Когда птицы раскрывают клюв, можно увидеть язык, зубов у них нет, это облегчает массу их черепа.
Рисунок 30.1 – Внешнее строение голубя

568
По бокам головы располагаются большие глаза, которые, как и у пресмыкающихся, защищены верхними и нижними веками и полупрозрачной мигательной перепонкой. За глазами находятся ушные отверстия.
Голова расположена на длинной и подвижной шее. Она позволяет птице, не меняя положения туловища, собирать корм, оглядываться по сторонам, чистить перья на всем теле.
Кожные покровы и их производные. Кожа птиц тонкая, не имеет каких-
либо костных образований и почти лишена желез (исключение – копчиковая железа, секрет которой служит для смазывания перьев и для придания перьевому покрову водонепроницаемости, сильно развита у водоплавающих птиц). Обильны и разнообразны роговые производные эпидермиса: верхняя и нижняя челюсти покрыты роговыми чехлами, образующими клюв; на концах пальцев – когти; на нижней части ног – роговые щитки.
Тело покрыто перьями, которые располагаются не повсеместно, а лишь на некоторых участках – птерилиях, на других участках – аптериях – перьев вовсе или почти нет (рисунок 30.2).
слева – вид со спинной стороны, справа – с брюшной. Птерилии: 1 – головная; 2 – спинная; 3 – крыловая; 4 – плечевая; 5 – бедренная; 6 – ножная; 7 – хвостовая; 8 –
грудная. Аптерии: 9 – верхняя крыловая; 10 – боковая; 11 – шейная; 12 – нижняякрыловая; 13 – брюшная; 14 – ножная
Рисунок 30.2 – Расположение птерилий и аптерий у кедровки
(Nucifraga caryocatactes)
Такое расположение перьев имеет приспособительное значение, так как облегчает сокращение мышц, подвижность кожи и перемещение перьев на туловище, связанное с движением крыльев.
Перья птиц различны по строению и функции. Контурные перья покрывают тело снаружи. Состоят из полого стержня, к которому прикреплены две боковые пластинки – опахала. Часть стержня, к которой прикреплены опахала, называют ствол, часть стержня, которая погружена в

569
кожу – очин (рисунок 30.3).
1 – опахало; 2 – стержень; 3 – обычная поверхность; 4 – пуховая часть; 5 – очин
Рисунок 30.3 – Строение пера
Опахало состоит из многочисленных бородок первого порядка, на которых сидят бородки второго порядка с мелкими крючочками, сцепляющими их между собой.
Контурные перья – основа оперенья (рисунок 30.4). В зависимости от расположения на теле различают: маховые – расположенные по заднему краю передних конечностей и формирующие лопасти крыльев; рулевые – длинные перья хвоста; кроющие – покрывающие крылья сверху и снизу; надхвостье – верхняя часть хвоста и т. д.
1 – маховые 1-го порядка; 2 – большие кроющие; 3 – крылышко; 4, 8 – маховые 2-го порядка; 5 – оперение плеча; 6 – средние кроющие; 7 – кроющие крыла
Рисунок 30.4 – Перья крыла птицы
Основная роль: предохраняют тело от потери тепла и механических воздействий; образуют гребную лопасть крыла и рулевую плоскость хвоста. Их число от 100 у колибри до 25 тыс. у лебедей.
Пуховые перья. Мелкие, находятся под контурными. Стержень их тонок, бородок второго порядка нет, поэтому опахала не образуют сомкнутых пластинок. Часто стержень пера настолько укорочен, что бородки отходят от вершины одним пучком. Такое перо называют
570
собственно пухом. Развиты сильно у водоплавающих и видов, обитающих в холодных странах. Основная роль – уменьшение теплоотдачи.
Нитевидные перья. Находятся среди пуха. В углах рта у многих птиц имеются щетинки.
Перья регулярно сменяются. У многих птиц в году бывает не одна, а две или три линьки (меняется обычно не все оперение, а только определенные его участки). Оперение птицы должно периодически сменяться для того, чтобы выполнять свойственные ему функции. Во время процесса линьки у птиц происходит замена старого перьевого покрова на новый. Этот процесс жизненно важен для птиц. Ведь перья со временем просто изнашиваются, а также теряют свои теплозащитные свойства и отражаются на способности летать. Обновление оперения у птиц приурочено к тому времени, когда птица свободна от забот, связанных с размножением. Токовое поведение, откладка яиц, насиживание и, наконец, выкармливание выводка – все это требует больших энергетических затрат. Когда выводок распался, для многих птиц перед отлетом в южные края наступает самое подходящее время сменить старое перо на новое. Линька у разных видов птиц происходит по–разному. Куриные птицы линяют не сразу, а постепенно. Совсем медленно линяют хищные птицы. Им приходится сменять перо исподволь, чтобы летные качества птицы не ослабевали. А вот птицы, связанные с водой, и прежде всего пластинчатоклювые (гуси, лебеди, утки), чистиковые птицы, поганки, гагары и ряд других видов, которые могут прокормиться и без помощи полета, линяют бурными темпами. Они забираются в малодоступные места, в "крепи", быстро теряют там маховые и рулевые перья и на некоторое время становятся совершенно беспомощными, так как в случае опасности не могут взлететь. Кряковая утка становится нелетающей птицей на 20–25 дней. По некоторым наблюдениям, лебеди не летают 40–50 дней. При линьке происходит не только смена оперенья, но меняется и его структура (больше перьев в зимнем оперении, зимние перья длиннее и т.д.).
Мускулатура более дифференцирована, чем у рептилий, что обусловлено сложными движениями при полете, хождении, лазании, добыче пищи. Наиболее крупные мышцы, приводящие в движение крылья, располагаются на туловище, а к самим конечностям идут сухожилия. Основная масса мускулатуры располагается не на спинной стороне тела, а на груди, где лежат мышцы, приводящие в движение крылья.
Особого внимания заслуживают мышцы конечностей. Грудные мышц, прикрепляющиеся к килю грудины и достигающие 20% общей массы тела птиц, служат для опускания крыл. Лежащие под нимиподключичные мышцы имеют меньшие размеры и служат для поднятия крыльев.
Очень сложную мускулатуру имеют задние конечности (до 35 мышц). Особый интерес представляет имеющаяся у некоторых видов обходящая мышца ноги. Начинается она на тазе, тянется вдоль бедра, далее в виде тяжа перекидывается через колено и затем соединяется со сгибателем пальцев.