

Osnovy_zoologii
.pdf481
самостоятельный класс панцирных рыб (Aphetohyoidea). К ним же причисляют и палеоспондилид (Palaeospondiloidei), ранее относимых к ископаемым бесчелюстным.
Хрящевые рыбы (Chondrichthyes) обособились от панцирных рыб еще в девоне (рисунок 24.2). Они характеризовались тем, что подъязычная дуга у них потеряла дыхательную функцию и превратилась в подвесок (hyomandibulare) челюстного аппарата, в то время как у панцирных рыб гиоидная дуга сохраняла функцию жаберной дуги. Хрящевые рыбы не имели наружного костного панциря и вели подвижный нектонный образ жизни, став активными хищниками морей. К ним относятся брадиодонты (Bradiodonti) и, по-видимому, их современные потомки химеровые (Holocephala), современные акуловые рыбы – эласмобранхии (Elasmobranchii), ископаемые акуловые рыбы плейракантоды (Pleuracanthodii) и кладоселахии (Cladoselachii). Все эти хрящевые рыбы господствовали в морях палеозойской эры, и в каменноугольный период они вытеснили агнат. Однако в пермский период они сами уступили место более совершенным костным рыбам.
Из всей многообразной фауны хрящевых рыб палеозойской эры в последующие эпохи сохранились лишь две их ветви – современные эласмо- бранхии и химеровые. Эласмобранхии сохранились потому, что у них выработался в процессе эволюции ряд прогрессивных черт организации (высокоразвитые органы чувств и головной мозг, внутреннее оплодотворение и откладка крупных, одетых скорлупой яиц), позволявших им конкурировать с высшими рыбами. Химеровые рыбы ушли от конкуренции, приспособившись к жизни в глубоких слоях моря.
Костные рыбы (Osteichthyes) ответвились от общего ствола рыб чрезвычайно рано (рисунок 24.3). Их ископаемые остатки встречаются почти одновременно с остатками древних хрящевых рыб уже в пресноводных отложениях девона.
Костные рыбы характеризуются следующими прогрессивными особенностями строения, давшими им преимущество по сравнению с древними хрящевыми рыбами. Их более легкий и прочный костный скелет давал преимущество в движении, так же как и новый важный орган – плавательный пузырь. У них образовалась жаберная крышка и более совершенный способ дыхания. Первичные костные рыбы уже в девоне распались на две группы.
Одна группа – лучеперые, или лучеплавниковые (Actinopterygii), эволюционировала в сторону приспособления к жизни в открытых водоемах и явилась родоначальником высших костных рыб; хрящевых ганоидов, или осетровых (Chondrostei), палеонисцид (Palaeoniscoidea), костных ганоидов (Holstei) и костистых рыб (Teleostei). Они характеризуются присутствием дорзального плавательного пузыря, служащего важным приспособлением при плавании, высокоразвитыми органами зрения и мозжечком головного мозга. Наиболее совершенная в этом отношении самая молодая группа

482
костистых рыб (Teleostei) появилась лишь в конце триаса и достигла очень быстро полного господства во всех водоемах земного шара как морских, так и пресноводных (из 20 тыс. видов всех рыб около 19,5 тыс. принадлежат костистым рыбам).
Рисунок 24.2 – Филогенетическое древо |
Рисунок 24.3 – Филогенетическое древо |
панцирных и хрящевых рыб |
костных рыб |
Другая группа древнейших костных рыб заселяла прибрежные зоны пресноводных водоемов и характеризовалась приспособлением к жизни в прибрежных зарослях. Плавники их приспособлены к поддержанию тела на дне, имеется тяжелый чешуйчатый покров из сложных ганоидных или космоидных чешуй. Органы зрения и мозжечок развиты слабо. В дополнение к жабрам у них развились брюшные плавательные пузыри как дополнительные органы дыхания. В черепе обнаружено присутствие хоан, указывающих на наличие легочного дыхания, поэтому их в настоящее время объединяют в общий подкласс хоановых (Choanichthyes, или Sarcopterygii).
В эту группу рыб относят кистеперых (Crossopterygii) и двоякодышащих (Dipnoi). Из кистеперых одна группа (Rhipidistia) вымерла еще в пермский период, а другая (Coelacantini) в виде реликтов дошла до настоящего времени (Latimeria). Двоякодышащие рыбы ответвились от общих предков с кистеперыми еще в девоне и в виде реликтов продолжают существовать и поныне (Ceratodus – в Австралии, Lepidosiren– в Америке и
Protopterus– в Африке).
Весьма близок по образу жизни к кистеперым рыбам африканский многопер, или полиптерус (Polypterus). Однако многие ихтиологи считают это сходство конвергентным и по наличию у полиптеруса ганоидных чешуй считают многопера родственным ископаемым палеонисцидам. Вместе с
483
хрящевыми ганоидами их относят в группу древних лучеперых рыб (Paleopterygii). В последнее время многоперовых выделяют в самостоятельный подкласс Brachiopterygii.
По палеонтологическим данным, древняя группа кистеперых (Rhipidistia) явилась родоначальником для предков наземных позвоночных, и ветвь первичных четвероногих (Quadrupeda) отводят в девонском периоде от этой группы рыб. Они имели мясистые плавники, приспособленные к ползанию по дну, и строение черепа, очень сходное с черепом древнейших ископаемых амфибий – стегоцефалов.
Рыбы возникли в пресных водоемах и только в последующем, хотя и довольно быстро, заселили Мировой океан.
484
ТЕМА 25: ЭКОЛОГИЯ РЫБ
ПЛАН:
25.1Условия жизни рыб в водной среде, лимитирующие факторы
25.2Жизненный цикл рыб.Миграции
25.3Ориентация и поведение рыб
25.4Роль рыб в водных экосистемах и значение для человека
25.1 Условия жизни рыб в водной среде, лимитирующие факторы
Внешнюю среду, рассматриваемую как фактор, влияющий на рыб, разделяют на биотическую и абиотическую.
Биотическая среда – это животный и растительный мир организмов, окружающих и связанных с рыбой. Влияние биотической среды на рыб имеет огромное значение. Это влияние осуществляется различными путями: путем конкуренции в пище между отдельными видами рыб, взаимоотношений между хищником и жертвой, влияний организмов на режим водоемов.
Абиотическая среда – это физические и химические свойства среды обитания рыб. К наиболее важным абиотическим факторам, влияющим на жизнедеятельность рыб, относятся соленость, температура, содержание газов в воде и др.
Температура воды. Рыбы могут жить при самой разнообразной температуре. Так, рыбка лукания живет и размножается в источниках горной Калифорнии, где температура воды выше 50°С. С другой стороны, даллия и караси выдерживают замерзание зимой, находясь в спячке на дне водоемов. Однако кровь и жабры у них не промерзают. При их промерзании гибель неизбежна. По отношению к температурному фактору рыб делят на теплолюбивых (осетровые, гартовые, сомовые, угревые и др.) и холодолюбивых (тресковые, лососевые и др.). Однако не менее важна амплитуда температур, при которых рыбы могут жить. Амплитуда температур для каждого вида рыб может быть очень разнообразной. Одни рыбы могут жить в пределах изменения температур в 5–7°С, другие в десятки градусов. Поэтому различают рыб эвритермных и стенотермных.
Эвритермные– такие рыбы, которые обитают при широких амплитудах температур, например, щука, плотва, окунь, маринка. Они могут обитать в пределах температуры от 3 до 25°С.
Стенотермные– рыбы, обитающие при узких температурных амплитудах. Они представлены главным образом рыбами типично арктическими, типично тропическими и глубоководными, которые плохо переносят даже незначительное изменение температуры.
Температура имеет влияние на ход жизненных процессов: сроки икрометания, развитие икры, скорость роста, газообмен. Количество потребляемого кислорода с повышением температуры увеличивается и,
485
наоборот, уменьшается с понижением температуры.
Температура влияет на питание рыб. Каждый вид наиболее активно питается при определенном интервале температур. Оптимум питания форели наблюдается при 10–12°С, щуки при 15–16°C, сазана при 20–28°С, а арктическая рыба сайка активнее всего питается при температуре минус 2°С. Выше и ниже определенной температуры рыбы вообще прекращают питаться. Так, ручьевая форель не питается при температуре воды ниже 3°С и выше 18°С.
Поскольку рыба приспособлена к жизни при определенной температуре, ее распределение в водоеме обычно связано с распределением температур. При разведке промысловых концентраций рыб часто ориентируются на распределение температур в водоеме. В Карское море в период потеплений проникает треска. Наоборот, в периоды похолоданий арктические виды спускаются в более низкие широты. Так, в Белое море в массовом количестве заходит сайка.
Соленость. Рыбы приспособились к жизни в воде различной солености. В пресной воде солей не более десятых долей грамма на 1 л (в основном углекислые и сернокислые). Нормальная морская вода содержит 35% (35 г соли на 1000 г воды) солей, преимущественно хлористых.
По отношению к амплитуде солености рыб делят на эвригалинных, способных переносить большие колебания солености, и стеногалинных, способных переносить лишь незначительные колебания солености. Примером эвригалинных рыб может служить каспийская рыба-игла, некоторые бычки, примером стеногалинных рыб – некоторые рыбы коралловых рифов.
Соленость играет в жизни рыб большую роль. Плотность воды зависит от количества растворенных солей, следовательно, условия плавания зависят от солености. Газовый режим также зависит от солености. Чем больше растворено в воде солей (при одной и той же температуре и одинаковых условиях), тем меньше в ней кислорода.
По месту обитания рыб подразделяют на четыре экологические группы: морских, пресноводных, проходных и солоновато-водных. Рыба, перенесенная из одной среды в другую, может погибнуть из-за большой разницы осмотического давления в органах и клетках тела и давления наружной среды. У проходных рыб при переходе из морской воды в пресную, происходит перестройка осморегуляторного аппарата, поэтому они задерживаются на некоторое время на границе соленых и пресных вод.
Растворенные в воде газы.В воде в больших количествах растворены кислород и азот, в значительно меньших – углекислый газ. Распределение газов в атмосферном воздухе и в воде неодинаково. Соотношение кислорода и азота в воздухе равно примерно 1:4, а в воде 1:2. Однако абсолютное количество кислорода в единице объема больше в воздухе, чем в воде. Так, например, при температуре 20°С в 1 л воздуха в 33 раза больше кислорода, чем в 1 л воды. В этом отношении вода представляет худшую
486
среду для дыхания, чем воздух.
Насыщение воды кислородом происходит из воздуха. Насыщение глубинной воды природных водоемов кислородом происходит за счет перемешивания воды ветрами и циркуляции из-за колебаний температуры. С увеличением температуры растворимость газа в воде уменьшается. В морях и океанах газовый режим относительно постоянен, так что рыбы здесь находятся в более благоприятных условиях, чем в пресноводных водоемах.
Рыбы по-разному реагируют на количество растворенного в воде кислорода. Одни нуждаются в очень высоком его содержании (лосось, сиг, форель, судак), другие менее требовательны (плотва, окунь, щука), третьи удовлетворяются совершенно ничтожным количеством (карась, линь). Для каждого вида рыб существует как бы определенный порог содержания кислорода в воде, ниже которого они становятся вялыми, плохо питаются и, наконец, погибают.
Одни рыбы (кумжа, гольцы) не терпят даже малейшего кислородного голодания и населяют водоемы только с прозрачной, холодной, богатой кислородом водой. Другие рыбы (караси) живут даже в болотах. Потребность в кислороде зависит от образа жизни. Малоподвижные донные рыбы потребляют меньше кислорода, чем пелагические, быстродвигающиеся.
Температура оказывает большое влияние на потребление рыбой кислорода. При повышении температуры количество потребляемого кислорода увеличивается.
25.2 Жизненный цикл рыб. Миграции
Жизненный цикл рыб складывается из ряда последовательно сменяющих друг друга этапов: созревания, размножения, нагула, зимовки. Во время каждого из этапов жизненного цикла рыбы нуждаются в специфических условиях среды, которые они находят в различных, зачастую далеко отстоящих друг от друга местах водоема, а иногда и в разных водоемах.
Так, условия, необходимые для нереста, отличны от условий, обеспечивающих наилучший нагул рыб. Своеобразные требования предъявляют рыбы к условиям зимовки и т.д. Это приводит к тому, что в поисках мест, подходящих для каждого жизненного отправления, рыбы совершают более или менее значительные перемещения. У видов, населяющих мелкие замкнутые водоемы (пруды, озера) или реки, перемещения имеют ничтожные масштабы, хотя и в этом случае они все же выражены вполне отчетливо. Выделяют нерестовые, кормовые,
зимовальные миграции.
Миграции рыб.Миграциями называются закономерные перемещения рыб по определенным путям в определенное время года. Эти закономерные странствования представляют собой одно из интереснейших явлений в
487
жизни рыб и имеют чрезвычайно важное значение для их промысла. Мигрировать могут отдельные особи, небольшие группы рыб и
обширные стада или косяки. Редкие рыбы ведут малоподвижный образ жизни, обитая все время в определенном небольшом районе. Большинство рыб предпринимают более или менее протяженные перемещения, собираясь для этого иногда в очень большие стаи или косяки. Эти косяки облавливаются рыбаками определенных районов, в которые рыба приходит или через которые она проходит. Пути миграций рыб не всегда остаются неизменными. Иногда в связи с изменением внешних условий меняются и миграционные пути рыб. Миграции могут распределяться на пассивные и активные.
Пассивные миграции – это перенос течением личинок или икринок рыб. Примером пассивных миграций может служить снос выклюнувшихся личинок мурманской сельди струями Нордкапского течения Гольфстрима от мест нереста у северо-западного побережья Норвегии в Баренцево море.
Активно мигрируют взрослые рыбы. Среди активных миграций различают кормовые, нерестовые и зимовальные.
Миграции делятся на анадромные и катодромные. Анадромными миграциями называют такие перемещения, при которых рыбы движутся вверх по реке или из морских просторов к берегам. Катодромные миграции – это движение рыб вниз по реке или от берегов в открытое море.Например, миграции из моря дальневосточных лососей для нереста в верховья рек или каспийских сельдей и осетровых в Волгу, так же как трески из открытого моря к побережью, являются анадромными, а выход для нереста речного угря из рек в океан или отход морской камбалы в глубокие соленые участки моря
– катадромными. Приведем примеры, характеризующие отдельные типы миграций.
Кормовые миграции. Примеров активных миграций с целью питания очень много. Черноморские кефали, морские рыбы в молодом возрасте для питания растительными остатками (детритом) весной заходят в мелкие солоноватоводные прибрежные озера– лиманы. Они проходят в них через узкие, иногда специально вырытые каналы. С наступлением похолодания откормившаяся и достигшая промысловых размеров кефаль уходит обратно в море. Или, например, миграция трески Баренцева моря (рисунок 25.1). Эта треска после размножения у берегов Норвегии около Лофотенских островов, исхудалая, тощая, голодная, движется, питаясь, в Баренцево море. Питается треска в Баренцевом море, главным образом, рыбой: сельдью, мойвой и в меньшей степени ракообразными. Можно указать целый ряд других морских рыб, совершающих обширные миграции с целью питания. Особенно значительны передвижения тунцов, меч-рыбы и некоторых акул, переходящих из одних океанов в другие в поисках пищи.
Нерестовые миграции. Особенно интересны и наиболее изучены нерестовые миграции проходных рыб. Типичным примером является миграция атлантического лосося (семги). Атлантический лосось для
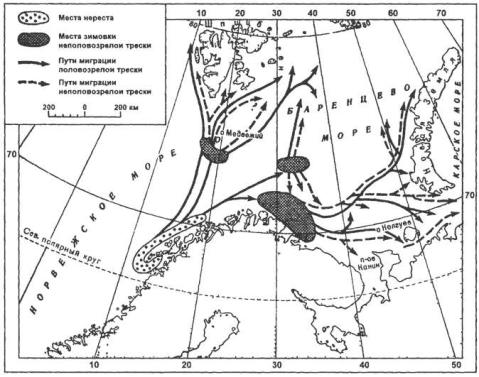
488
размножения из северных морей, где он обитает, входит в реки и, как правило, высоко по ним поднимается. Таким образом, его нерестовые миграции являются примером анадромных миграций. Атлантические лососи мечут икру осенью. В реки они входят в различные периоды года, в разном возрасте и различных размеров. Стремление к местам размножения проявляется у лосося очень сильно. Он преодолевает на своем пути вверх по реке маленькие пороги и даже водопады, совершая для этого иногда громадные прыжки до 4 м над водой. Нерест лосося происходит в мелких местах верховьев реки с ровным песчаным галечным дном, с прозрачной и холодной водой. Другой пример нерестовых анадромных миграций проходных рыб – миграции осетровых рыб (шип, осетр, севрюга и белуга). Осетровые начинают нерестовую миграцию весной. Высота подъема их по реке зависит от наличия удобных для икрометания мест с галечным дном.
Рисунок 25.1 – Схема миграций взрослой трески
Морская норвежская сельдь перед размножением нагуливается далеко к северо-западу от Скандинавии, у Фарерских островов, и даже в водах у Шпицбергена. В конце зимы косяки сельди начинают двигаться к берегам Норвегии и достигают их в феврале-марте. Нерест происходит в фьордах близ берега на мелких местах. Выведшиеся личинки лишь отчасти остаются в фьордах, большая же их масса увлекается северо-восточной ветвью Гольфстрима – Нордкапским течением вдоль берегов Скандинавии на север. Такую пассивную миграцию личинки начинают зачастую в еще очень раннем возрасте, когда у них сохраняется желточный пузырь. За 3-4 месяца,

489
до конца июля - начала августа, они проделывают путь в 1000-1200 км и достигают берегов Северной Норвегии (рисунок 25.2).
Обратный путь молодые сельди проходят активно, но медленно – за 4-5 лет. Они продвигаются на юг этапами ежегодно, то приближаясь к берегам, то отходя в открытое море. В четырех- или пятилетнем возрасте сельди становятся половозрелыми и достигают к этому времени района икрометания
– мест, где родились. Этим заканчивается первый, "юношеский", этап их жизни – период далекого путешествия на север. Второй период – период зрелости – связан с ежегодными миграциями от места нагула к местам нереста и обратно.
Рисунок 25.2 – Схема миграции норвежской сельди
Значительно реже среди проходных рыб наблюдаются примеры катодромных миграций, когда реки являются для рыбы местом питания, а море – местом размножения. Такой тип миграции встречается у европейских речных угрей (рисунок 25.3). Европейский угорь по достижении половой зрелости начинает уходить из рек, где он кормился и рос, в море. В море угорь движется от берегов Европы к берегам Центральной Америки, где над большими глубинами и находятся места его размножения. Морские рыбы также совершают нерестовые миграции. Атлантическая треска, например, совершает по окончании нагула нерестовую миграцию из восточной части Баренцева моря к Лофотенским островам. Морская камбала обычно отходит для икрометания от берега на значительные глубины.
Пресноводные рыбы также для размножения поднимаются выше по рекам или идут из озер в реки. Таковы стерлядь, волховской сиг, налим, жерех. Однако обычно пути нерестовых миграций пресноводных рыб короче, чем проходных или морских.

490
Зимовальные миграции. Типичны зимовальные миграции рыб, обитающих в Северном Каспии. Сазан, лещ, судак, собираются осенью в большие стаи и мигрируют к берегам и в дельту рек, главным образом в Волгу. Здесь эти рыбы залегают в особых ямах на зиму. Важное значение для промысла имеют зимовальные миграции азовской хамсы осенью из Азовского моря через Керченский пролив в Черное море. Хамса – типично пелагическая планктоноядная рыба. После откорма в Азовском море она собирается осенью в большие стаи и выходит через Керченский пролив в Черное море. Затем она движется к южным берегам Кавказа, опускается зимой с поверхности на глубину, где и зимует. Весной, поднявшись к поверхности, хамса мигрирует обратно в Азовское море для нереста и нагула.
Рисунок 25.3 – Карта распространения личинок европейского речного угря в Атлантическом океане
Густо заштрихованное место – область икрометания европейского угря в Саргассовом море. Цифры обозначают среднюю длину личинок угря в мм. Линиями покрыта область Атлантического океана, где встречаются личинки угря; за ее границей к востоку встречаются угри на стадии метаморфоза. Черная полоса, окаймляющая берега,– область распространения угря в реках
Приведенные типы миграций нельзя рассматривать изолированно. Миграции у каждой рыбы связаны одна с другой, переводят одна в другую. Все типы миграций у рыб представляют собой отдельные этапы общего миграционного годового цикла.
Миграции могут происходить пассивно когда мигранты без