

Osnovy_zoologii
.pdf331
форм с удлиненной раковиной в форме буравчика со множеством оборотов спирали. Особую группу образуют кораллобионты, у которых раковина в форме извитой трубки прирастает к скелетам коралловых полипов. Верметиды, которые также имеют трубковидную раковину, образуют плотные поселения на твердом субстрате. Приспособились к плавающему образу жизни киленогие и крылоногие моллюски, относящиеся к разным подклассам. Также параллельно, независимо друг от друга, совершили выход на сушу некоторые переднежаберные и легочные; при этом они стали развиваться без метаморфоза и приобрели приспособления к воздушному дыханию. Наиболее специализированными среди них оказались легочные, которые распространились почти по всем ландшафтам суши и различным ярусам биоценозов (от почвенного до древесного). Часть сухопутных моллюсков совершили эволюционный возврат в водную среду – с образованием вторично-водных пресноводных форм с легочным дыханием.
16.3 Своеобразие в строении, физиологии, размножении и развитии двустворчатых моллюсков
К классу двустворчатых относятся исключительно водные, малоподвижные донные моллюски с двустворчатой раковиной, полностью прикрывающей их тело. Класс насчитывает более 20 тыс. видов. Они способны к агрегации и образуют массовые скопления. Двустворчатые в основном относятся к группе биофильтраторов, питающихся взвешенными в воде частицами органических веществ и мелким планктоном, и потому играют существенную роль в биологической очистке вод. У большинства видов сильно развиты пластинчатые жабры, выполняющие не только дыхательную, но и фильтрующую функцию. В связи с пассивным движением и питанием у двустворчатых редуцировалась голова. Все особенности их внешнего и внутреннегостроения отражают их экологическую специализацию к малоподвижному или неподвижному образу жизни.
Внешнее строение. Двустворчатые характеризуются наличием раковины из двух створок, клиновидной роющей ногой и отсутствием головы. У неподвижных форм нога, как правило, редуцирована.
Форма и размеры раковин двустворок сильно варьируют. Раковины самых мелких двустворчатых не превыша ют 2–3 мм в длину. Гигант среди моллюсков – тридакна, обитающая в тропических морях, может достигать 1,4 м в длину, а ее масса может быть до 200 кг. Створки раковины чаще симметричны, как у сердцевидок и беззубок. У некоторых видов может наблюдаться асимметрия створок раковины.
Стенки раковины обычно состоят из трех слоев: наружного – конхиолинового (периостракум), внутреннего – известкового (остракум) и нижнего – перламутрового (гипостракум). Раковина выделяется мантией. Прирост раковины осуществляется краем мантии. На раковине видны
332
концентрические линии, отражающие ее неравномерный рост в изменяю- щихся условиях среды. Конхиолиновый слой имеет разнообразную покровительственную окраску. На вершинах створок этот слой часто стирается. Внутренний перламутровый слой состоит из мелких плоских известковых пластиночек, соединенных конхиолином. Такое строение перламутра вызывает интерференцию света, вследствие чего перламутровый слой переливает всеми цветами радуги. Если между мантией и створкой раковины попадает какая-нибудь инородная частица, то она обволакивается концентрическими слоями перламутра и образуется жемчужина. Слой перламутра утолщается с возрастом моллюска и ростом его раковины.
Створки раковины соединяются на спинной стороне связкой – лигаментом, состоящим из утолщенного рогового слоя раковины. У большинства видов на створках раковины имеется замок. Это зубцы и углубления на спинном крае раковины с внутренней стороны . Зубцы замка одной створки входят в соответствующие углубления другой и тем самым препятствуют смещению створок. Замок может быть равнозубый (таксодонтный) или разнозубый (гетеродонтный). У некоторых двустворок замок редуцирован (беззубка – Anodonta). Створки раковины живого моллюска могут раскрываться и закрываться с помощью мускулов- замыкателей (один или два). Это толстые пучки мышц, соединяющие обе створки. При их сокращении створки закрываются, при расслаблении открываются. Механизму раскрывания створок способствует лигамент, который в положении закрытой раковины находится в натянутом состоянии, подобно пружине, а при расслаблении мускулов-замыкателей приходит в исходное положение, раскрывая створки. В некоторых случаях раковина может быть редуцированной. Так, у моллюска-древоточца (Teredo) раковина прикрывает лишь 1/20 тела и представляет собой сверлящий аппарат.
Нога клиновидной формы служит для зарывания в грунт и медленного ползания. У некоторых прикрепляющихся к субстрату форм в ноге имеется особая биссусная железа, выделяющая биссусные нити, при помощи которых они как бы прирастают к твердой поверхности дна (мидия). У многих неподвижных форм нога редуцируется полностью (устрица).
Мантия и мантийная полость. Мантия имеет форму двух складок кожи, свешивающихся со спины по бокам к брюшной стороне. Наружный железистый слой мантии выделяет раковину. Внутренняя поверхность мантии покрыта мерцательным эпителием, движение ресничек которого обеспечивает ток воды в мантийной полости. Снизу складки мантии могут быть свободными, как у беззубки, а могут срастаться, образуя лишь отверстия для ноги в передней части и задние отверстия сифонов.
У роющих форм сифоны, образованные мантией, длинные, в виде двух трубок, выступающих из грунта. По нижнему, вводному сифону вода

333
входит в мантийную полость, а по верхнему, выводному сифону выходит. Вода приносит в мантийную полость моллюска пищевые частицы и кислород.
В мантийный комплекс органов входят: нога, две жабры, две ротовые лопасти, осфрадии и отверстия пищеварительной, половой и выделительной систем органов.
Пищеварительная система двустворчатых отличается своеобразием в связи с пассивным способом питания путем фильтраций (рисунок 16.6). У них имеется фильтрующий аппарат. Вода, поступающая через вводный сифон в мантийную полость, направляется к переднему концу тела, омывая жабры и ротовые лопасти. Движение воды в мантийной полости обеспечивается мерцательным эпителием, покрывающим жабры, ротовые лопасти и внутреннюю поверхность мантии. На жабрах и ротовых лопастях имеются рецепторные клетки (органы вкуса) и ресничные желобки, по которым мелкие частицы пищи, отсортированные от минеральных частиц, транспортируются в рот. Рот находится в передней части тела около переднего мускула-замыкателя. Изо рта пища поступает в пищевод, затем в энтодермальный желудок. Глотка, радула и слюнные железы в связи с редукцией головы у двустворчатых отсутст вуют.
1 – линия, по которой обрезана мантия;2– мускул-замыкатель; 3 –рот; 4– нога; 5–ротовые лопости; 6,7–жабры; 8–правая мантия; 9– вводной сифон; 10– выводной сифон; 11– задняя кишка; 12–перикард
Рисунок 16.6 – Схема строения двустворчатого моллюска
В желудок впадают протоки двулопастной печени. Кроме того, в желудке имеется кристаллический стебелек, выделяющий пищеварительные ферменты. От желудка отходит средняя кишка, которая потом переходит в заднюю, открывающуюся анальным отверстием в мантийную полость. Задняя кишка двустворчатых обычно пронизывает желудочек сердца. Экскременты из мантийной полости током воды
334
выбрасываются через выводной сифон наружу.
Нервная система двустворчатых упрощена в сравнении с брюхоногими. В связи с редукцией глотки церебральные ганглии слились с плевральными и образовались цереброплевральные сдвоенные узлы. В ноге имеются педальные ганглии, связанные коннективами с первой парой узлов. В заднем конце тела под задним мускулом-замыкателем располагается третья пара узлов – висцеропариетальные, которые иннервируют внутренние органы, жабры и осфрадии.
Органы чувств развиты слабо. В ноге имеются статоцисты – органы равновесия, иннервируемые церебральными ганглиями. У основания жабер находятся осфрадии – органы химического чувства. Рецепторные клетки располагаются на жабрах, ротовых лопастях, краях мантии и на сифонах. Имеются случаи образования глаз по краю мантии у гребешка или на сифонах у сердцевидок.
Органы дыхания представлены ктенидиями – жабрами. В пределах класса двустворчатых жаберный аппарат варьирует. У наиболее примитивных двустворчатых – первичножаберных (Рrоtobranchia) имеется пара типичных ктенидиев с перистыми лепестками. У нитежаберных (Filibranchia) жабры могут быть нитевидные или пластинчатые. Нитевидные жабры характеризуются тем, что их жаберные лепестки представлены длинными нитями, которые ниспадают в нижнюю часть мантийной полости, а затем загибаются вверх. При этом соседние нити скрепляются друг с другом с помощью жестких ресничек, образуя пластинки. Нитевидные жабры свойственны мидиям, устрицам, гребешкам. Еще более сложное строение у двустворчатых имеют пластинчатые жабры. Они представляют дальнейшую модификацию нитевидных жабр. У них возникают поперечные перегородки между восходящими и нисходящими участками нитей и между соседними нитями. Это приводит к образованию жаберных пластинок. Каждая жабра состоит из двух пластинок – наружной и внутренней. Наружная полужабра прилегает к мантии, а внутренняя – к ноге. Такие жабры характерны для беззубок и перловиц. И наконец, у перегородчатожаберных (Septibranchia) жабры редуцированы и преобразованы в жаберную перегородку с порами. Перегородка отгораживает верхнюю часть мантийной полости, выполняющую дыхательную функцию. Стенки этой дыхательной полости имеют густую сеть кровеносных сосудов, где происходит газообмен.
Морфологический ряд жабр – от ктенидиев к нитевидным и пластинчатым жабрам – отражает основную эволюционную тенденцию в изменении органов дыхания у двустворчатых моллюсков.
Кровеносная система. Сердце двустворчатых расположено на спинной стороне и состоит из одного желудочка и двух предсердий. Сквозь желудочек сердца проходит задняя кишка. Это объясняется тем, что сердце закладывается в эмбриогенезе как парное по бокам кишки, а затем эти зачатки соединяются над и под кишкой. Парное происхождение сердца и
335
двустворчатых подтверждается фактом наличия двух сердец у моллюска
Area.
Кровь циркулирует по сосудам и лакунам. От желудочка отходят передняя и задняя аорты, ветвящиеся на артерии, из которых кровь переходит в лакуны. Венозная кровь от внутренних органов собирается в крупную продольную лакуну под сердцем. Из лакуны кровь переходит в приносящие жаберные сосуды.
Окисленная артериальная кровь из жабер по выно сящим сосудам возвращается в сердце. Частично кровь, минуя жабры, проходит через почки, освобождаясь от продуктов обмена, и вливается в выносящие жаберные сосуды, впадающие в предсердия.
Органы выделения – почки, типичные для всех моллюсков. Почки двустворчатых с железистыми стенками называют боянусовыми органами. Они имеют V-образную форму и одним концом открываются в перикард, а другим – в мантийную полость.
Дополнительно выделительную функцию выполняет стенка перикарда, видоизмененная в виде парных желез – кеберовых органов.
Половая система. Двустворчатые моллюски раздельнополы. Парные гонады залегают в передней части тела и в основании ноги. У некоторых видов нет половых протоков и половые клетки выходят из гонад через разрывы тканей в мантийную полость.
Но могут быть и парные половые протоки (яйцеводы или семяпроводы), открывающиеся половыми отверстиями в мантийную полость. Оплодотворение наружное. Мужские половые клетки из мантийной полости самцов выходят через сифон наружу, а затем током воды затягиваются в мантийную полость женских особей, где и происходит оплодотворение яиц.Такое оплодотворение возможно только при тесных скоплениях моллюсков.
Развитие. У большинства двустворчатых развитие с метаморфозом. Из оплодотворенных яиц развивается планктонная личинка – велигер, выполняющая расселительную функцию.Впоследствии велигер оседает на дно, прикрепляясь биссусной нитью, теряет парус и превращается во взрослого моллюска.
У некоторых пресноводных двустворчатых (беззубка, перловица) метаморфоз протекает с образованием особой личинки – глохидия. Глохидий имеет тонкостенную двустворчатую раковину с крючками на брюшном крае. Он плавает в воде, хлопая створками, приводимыми в движение мощным мускулом-замыкателем. С помощью биссусной нити глохидии прикрепляются к жабрам проплывающих рыб и затем зубцами раковины внедряются в их ткани. Их развитие протекает в коже рыб. Из глохидиев формируются маленькие моллюски, которые через разрыв опухолевых бугорков на коже рыбы выпадают на дно. Эктопаразитизм личинок двустворок на рыбах обеспечивает расселение моллюсков вверх и вниз по течению рек. В некоторых случаях у пресноводных двустворок
336
встречаются случаи прямого развития.
Классификация. Для пластинчатожаберных принимается несколько различных систем. Классификация этой группы может основываться на разных признаках – на особенностях строения замка, мускулов-замыкателей раковины, жабр. По последнему признаку пластинчатожаберных делят на 4 отряда.
Отряд Первичножаберные (Protobranchia) – небольшая группа наиболее примитивных пластинчатожаберных, для которых характерны типичные ктенидии, нога с подошвой, статоцист, плевральные ганглии обособленные от церебральных, отсутствие специальных половых протоков. Живут преимущественно в северных морях обычно мелкие формы. Представители: Joldiа, Nucula.
Отряд Нитежаберные (Filibranchia). Жаберные лепестки вытянуты в длинные, сложенные пополам нити. Представители: ноев ковчег (Arcanoae), мидия (Mytilus); гребешок (Pecten), устрица (Ostrea); морская жемчужница
(Pinctada margaritifera); Lithophaga,проделывающая ходы в известняковых породах.
Отряд Настоящие пластинчатожаберные (Eulamellibranchia). Жабры превращены в двойные решетчатые пластинки. Объединяет большинство пресноводных пластинчатожаберных (жемчужница –Margaritifera, перловица
– Unio, беззубка – Anodonta, дрейссена – Dreissena) и многие морские виды (сердцевидка – Cardium; корабельный червь, или шашень – Teredo и камнеточец Pholas способны проделывать ходы в дереве, в мягких горных породах; Tridacna – типичный обитатель коралловых рифов и др.).
Отряд Перегородчатожаберные (Septibranchia). К ним относятся небольшие морские, преимущественно глубоководные моллюски, с редуцированными жабрами. Их мантийная полость поделена на 2 части мускулистой поперечной перегородкой с отверстиями. Газообмен осуществляется в верхней части мантийной полости,
Филогенетические отношения и экологическая радиация. Среди современных двустворчатых наиболее примитивны первичножаберные (Protobranchia). У них проявляются признаки предков: узловая нервная система с неслившимися церебральными и плевральными ганглиями, рудимент плоской подошвы на клиновидной ноге, настоящие перистые ктенидии, парные зачатки сердца.
От древних первичножаберных ведут свое начало моллюски с нитевидными или пластинчатыми жабрами –видоизмененными ктенидиями. У большинства видов нога имеет биссусную железу, но нередко железа присутствует только на фазе личинки. У многих видов, перешедших к неподвижному образу жизни, нога редуцирована. Экологическая специализация шла по пути совершенствования прикрепленного или роющего образа жизни, а также изменения жаберного аппарата, выполняющего дыхательную, фильтрующую и даже половую функции (на жабрах происходит развитие молоди). Наиболее уклонившийся надотряд
337
представляют перегородчатожаберные (Septibranchia), у которых жабры редуцировались, а дыхательную функцию стали выполнять наджаберные полости.
Центральную группу двустворчатых представляют донные полуповерхностные и слабозарывающиеся формы, от которых прослеживаются два основных пути специализации – к прикрепленным фильтраторам (типа мидий, устриц) и к роющим формам с длинными сифонами, среди которых наиболее узкоспециализированными являются древоточцы и камнеточцы. Неподвижные формы проявляют особенно большое многообразие. Одни из них прикрепляются к грунту биссусом и расположены вниз брюшной стороной (мидия). Другие оседают на мягкий грунт вниз заостренной вершиной раковины, подобно пинне, а их створки торчат брюшной стороной вверх. Тяжелые тридакны лежат на спине, приоткрыв створки. Большинство неподвижных двустворок лежат либо на правой створке, как гребешок, либо на левой, как устрица. Таким образом, среди двустворок использованы все возможные способы прикрепления и положения тела на грунте в песке, которых они захватывают нитевидными головными щупальцами. У них отсутствуют жабры, сердце рудиментарное. У личинок лопатоногих раковина закладывается в виде тонкой пластинки, которая перегибается на спинной стороне, а ее свободные края постепенно смыкаются на брюшной. Онтогенез лопатоногих показывает на возможный путь образования в процессе эволюции трубчатой раковины.
16.4 Головоногие моллюски как высшие представители типа
Головоногие – самые высокоорганизованные моллюски. Их справедливо называют «приматами» моря среди беспозвоночных животных за совершенство их приспособлений к жизни в морской среде и сложность поведения. Это в основном крупные хищные морские животные, способные активно плавать в толще воды. К ним относятся кальмары, осьминоги, каракатицы, наутилусы. Их тело состоит из туловища и головы, а нога преобразована в щупальца, расположенные на голове вокруг рта, и особую двигательную воронку на брюшной стороне тела. Отсюда произошло название – головоногие.Доказано, что часть щупалец головоногих образуется за счет головных придатков.
У большинства современных головоногих раковина отсутствует или рудиментарна. Только у рода наутилус (Nautilus) имеется спирально закрученная раковина, разделенная на камеры.
К современным головоногим относится всего 650 видов. Это древняя группа моллюсков, известная с кембрия. Вымершие виды головоногих были преимущественно раковинными и имели наружную или внутреннюю раковину.
Для головоногих характерны многие прогрессивные черты организации в связи с активным образом жизни морских хищников. Вместе с тем у них сохраняются некоторые примитивные признаки,
338
свидетельствующие об их древнем происхождении.
Внешнее строение. Особенности внешнего строения головоногих разнообразны в связи с различным образом жизни. Их размеры колеблются от нескольких сантиметров до 18 м у некоторых кальмаров. Нектонные головоногие обычно торпедовидной формы (большинство кальмаров), бентосные имеют мешковидную форму тела (многие осьминоги), нектобентосные – уплощенную (каракатицы). Планктонные виды, мелкие по размерам, имеют студенистое плавучее тело. Форма тела у планктонных головоногих может быть узкой или похожей на медуз, а иногда шаровидной (кальмары, осьминоги). Бентопелагические головоногие обладают раковиной, поделенной на камеры.
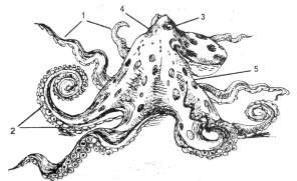
Тело головоногих состоит из головы и туловища (рисунок 16.7). Нога модифицирована в щупальца и воронку. На голове расположен рот, окруженный щупальцами, и крупные глаза. Щупальца образованы головными придатками и ногой. Это органы захвата пищи. У примитивного головоногого – кораблика (Nautilus) щупалец около 90; они гладкие, червеобразные. У высших головоногих щупальца длинные, с мощной мускулатурой и несут крупные присоски на внутренней поверхности. Число щупалец 8–10. У головоногих с 10 щупальцами 2 щупальца –ловчие, более длинные, с присосками на расширенных концах, а остальные 8 щупалец более короткие (кальмар, каракатица). У осьминогов, обитающих на морском дне, 8 щупалец одинаковой длины. Они служат осьминогу не только для захвата пищи, но и для передвижения по дну. У самцов осьминогов одно щупальце видоизменено в половое (гектокотиль) и служит для переноса половых продуктов в мантийную полость самки.
1 – щупальца; 2 – присоски; 3 – глаз; 4 – голова; 5 – туловище
Рисунок 16.7 – Внешнее строение осьминога
Воронка – производное ноги у головоногих, служит для «реактивного» способа движения (рисунок 16.8). Через воронку вода с силой выталкивается из мантийной полости моллюска, и его тело движется реактивно в противоположном направлении. У кораблика воронка не срослась на брюшной стороне и напоминает свернутую в трубку подошву ноги ползающих моллюсков. Доказательством того, что щупальца и воронка

339
головоногих –производные ноги, служит их иннервация от педальных ганглиев и эмбриональная закладка этих органов на брюшной стороне зародыша. Но, как уже отмечалось, часть щупалец головоногих – производные головных придатков.
Мантия на брюшной стороне образует как бы карман –мантийную полость, открывающуюся наружу поперечной щелью. Из этой щели выступает воронка. На внутренней поверхности мантии имеются хрящевые выступы –запонки, которые плотно входят в хрящевые углубления на теле моллюска, и мантия как бы пристегивается к телу.
Мантийная полость и воронка в совокупности обеспечивают реактивное движение. При расслаблении мускулатуры мантии вода входит через щель в мантийную полость, а при ее сокращении полость закрывается на запонки и вода выталкивается через воронку наружу. Воронка способна изгибаться вправо, влево и даже назад, что обеспечивает разное направление движения. Роль руля дополнительно выполняют щупальца и плавники – кожные складки туловища. Типы движения у головоногих разнообразны. Осьминоги чаще передвигаются на щупальцах и реже плавают. У каракатиц, кроме воронки, для движения служит круговой плавник. Некоторые глубоководные осьминоги зонтикообразной формы имеют перепонку между щупальцами – умбреллу и могут передвигаться за счет ее сокращений, подобно медузам.
1 – руки с присосками; 2 – ловчая рука; 3 – рот; 4 – отверстие воронки; 5 – воронка; 6 – хрящевые ямки запонок; 7 – анальный сосочек с анальным отверстием; 8 – почечные сосочки; 9 – непарный половой сосочек; 10 – жабры; 11 – плавник; 12 – линия отреза мантии; 13 – отогнутая мантия; 14 – хрящевые бугорки запонок; 15 – мантийный звездчатый ганглий
Рисунок 16.8 – Каракатица со вскрытой мантийной полостью (вид с брюшной
стороны)
Раковина у современных головоногих рудиментарна или отсутствует. У древних вымерших головоногих раковина была хорошо развита. Только
340
один современный род Nautilusсохранил развитую раковину.
У наутилуса спирально закрученная раковина разделена перегородками на камеры. Тело моллюска размещается только в последней камере, открывающейся устьем наружу. Остальные камеры заполняются газом, что обеспечивает плавучесть тела моллюска. Через отверстия в перегородках между камерами раковины проходит сифон – задний отросток тела. Клетки сифона способны выделять газы. При всплывании моллюск заполняет камеры раковины газом, а при опускании на дно камеры раковины заполняются жидкостью. Движителем у наутилуса является воронка, а раковина представляет собой гидростатический аппарат. Ископаемые наутилиды имели раковину, как у современного наутилуса. У полностью вымерших головоногих – аммонитов также была наружная, спирально закрученная раковина с камерами, но у них перегородки между камерами имели волнистое строение, что увеличивало прочность раковины. Именно поэтому аммониты могли достигать очень крупных размеров (до 2 м в диаметре). У другой группы вымерших головоногих – белемнитов (Belemnoidea) раковина была внутренняя, обросшая кожей. Белемниты по внешнему облику напоминали безраковинных кальмаров, но в их туловище находилась коническая раковина, поделенная на камеры. Вершина раковины заканчивалась острием – рострумом. Рострумы раковин белемнитов часто встречаются в меловых отложениях, и их называют «чертовыми пальцами». У некоторых современных безраковинных головоногих имеются рудименты внутренней раковины. Так, у каракатицы на спине под кожей сохраняется известковая пластинка, имеющая на срезе камерное строение. Только у спирулы (Spirula) под кожей находится вполне развитая спирально закрученная раковина, а у кальмара сохранилась от раковины лишь роговая пластинка. У самок современных головоногих – аргонавтов (Argonauta) развита выводковая камера, напоминающая по форме спиральную раковину. Но это лишь внешнее сходство. Выводковая камера выделяется эпителием щупалец, очень тонкая и предназначена для защиты развивающихся яиц.
Покровы. Кожа представлена однослойным эпителием и слоем соединительной ткани. В коже имеются пигментные клетки – хроматофоры. Головоногим свойственна способность быстро изменять окраску. Этот механизм контролируется нервной системой и осуществляется за счет изменения формы пигментных клеток. Так, например, каракатица, проплывая над песчаным грунтом, принимает светлую окраску, а над каменистым грунтом – темную. При этом в ее коже пигментные клетки с темным и светлым пигментом попеременно то сжимаются, то расширяются. Если перерезать зрительные нервы у моллюска, то он теряет способность изменять окраску. За счет соединительной ткани кожи образуются хрящи: в запонках, основаниях щупалец, вокруг мозга.
Головоногие, утратив в процессе эволюции раковину, приобрели другие защитные приспособления. Во-первых, многих из них спасает от