

Нейрохирургия
.pdfРис.
3.15. Сегментарная иннервация кожи туловища и конечностей ясничной и крестцовой сегментарных зон иннервации. Нарушение поверхностных
видов чувствительности при сохранных глубоких (или наоборот), свойственное очаговому поражению спинного мозга, обозначается как диссоциированный тип расстройств чувствительности.
При поражении задних канатиков имеется нарушение глубокой чувствительности ниже уровня поражения при сохранности поверхностной чувствительности (диссоциированный тип расстройств чувствительности), при этом наблюдается сенситивная атаксия. Движения становятся несоразмерными, неточными, при выполнении движений включаются мышцы, не имеющие прямого отношения к
выполняемому движению. При ходьбе больной чрезмерно разгибает ноги и выбрасывает их вперед, сильно топает («штампующая походка»). При включении зрения атаксия уменьшается. Атаксия в нижних конечностях выявляется при пяточно-коленной пробе, пробе Ромберга.
Поражение половины спинного мозга (синдром Броун-Секара) сопровождается снижением глубокой чувствительности и центральными двигательными расстройствами на стороне поражения и нарушением поверхностной чувствительности на противоположной. При полном поперечном поражении спинного мозга наблюдается проводниковый тип расстройства всех видов чувствительности ниже уровня поражения - параанестезия.
При поражении мозгового ствола возможен альтернирующий тип расстройства чувствительности: снижение поверхностной чувствительности в противоположных очагу конечностях (в результате поражения спинноталамического пути) и сегментарная гипестезия на лице на стороне очага (вследствие поражения ядра тройничного нерва).
Таламический тип расстройства чувствительности (при поражении таламуса) -
гемигипестезия в противоположных очагу конечностях на фоне гиперпатии, преобладание расстройств глубокой чувствительности, «таламические» боли (жгучие, периодически усиливающиеся и плохо купируемые анальгетиками). Если поражены чувствительные пути в задней ножке внутренней капсулы, выпадают все виды чувствительности на противоположной половине тела (гемигипестезия или гемианестезия). Как правило, одновременно имеется поражение и других проводящих путей (гемипарез, центральное поражение лицевого и подъязычного нервов, гемианопсия).
Корковый тип расстройства чувствительности (при поражении коры больших полушарий) проявляется парестезиями (покалывание, ползание мурашек, онемение) в половине верхней губы, языка, лица, в руке или ноге на противоположной стороне в зависимости от локализации поражения в постцентральной извилине. Парестезии могут иметь вид фокальных чувствительных эпилептических припадков (парциальные сенсорные припадки). Распознавание объектов при прикосновении (стереогноз) требует включения дополнительных ассоциативных полей коры, локализованных в теменной доле, где информация о размере, форме, физических свойствах (острота, мягкость, твердость, температура и др.) объекта интегрируется и может быть сопоставлена с теми тактильными ощущениями, которые имелись в прошлом. В связи с этим поражение нижней теменной дольки проявляется астереогнозом, т.е. утратой
способности узнавать предметы при прикосновении к ним на противоположной очагу стороне.
Нарушение мышечно-суставной чувствительности означает расстройство координации движений, неловкость при выполнении произвольных движений, гиперметрию. Это может проявляться в виде афферентного пареза, т.е.
расстройств двигательных функций, которые обусловлены нарушением мышечносуставного чувства при сохранной мышечной силе. Синдром афферентного пареза может быть одним из признаков поражения теменной доли.
ГЛАВА 4. ДВИЖЕНИЯ И ИХ РАССТРОЙСТВА
4.1. Пирамидная система Различают два основных вида движений - непроизвольные и произвольные. К
непроизвольным относятся простые автоматические движения, осуществляемые за счет сегментарного аппарата спинного мозга и мозгового ствола по типу простого рефлекторного акта. Произвольные целенаправленные движения - это акты двигательного поведения человека. Специальные произвольные движения (поведенческие, трудовые и др.) осуществляются при ведущем участии коры большого мозга, а также экстрапирамидной системы и сегментарного аппарата спинного мозга. У человека и высших животных осуществление произвольных движений связано с пирамидной системой, состоящей из двух нейронов - центрального и периферического.
Центральный мотонейрон. Произвольные движения мышц происходят в результате импульсов, идущих по длинным нервным волокнам из коры большого мозга к клеткам передних рогов спинного мозга. Эти волокна формируют двигательный (корково-спинномозговой), или пирамидный, путь.

Тела центральных мотонейронов расположены в прецентральной извилине в цитоархитектонических полях 4 и 6 (рис. 4.1). Эта узкая зона тянется вдоль центральной щели от латеральной (сильвиевой) борозды к передней части парацентральной дольки на медиальной поверхности полушария, параллельно чувствительной области коры постцентральной извилины. Подавляющее большинство мотонейронов залегает в 5-м кортикальном слое поля 4, хотя они встречаются и в соседних кортикальных полях. Преобладают малые пирамидные, или фузиформные (веретенообразные), клетки, дающие основу для 40% волокон пирамидного пути. Гигантские пирамидные клетки Беца имеют аксоны с толстой миелиновой оболочкой, обеспечивающие точные, хорошо скоординированные движения.
Нейроны, иннервирующие глотку и гортань, расположены в нижней части прецентральной извилины. Далее в восходящем порядке идут нейроны, иннервирующие лицо, руку, туловище, ногу. Таким образом, все участки тела человека спроецированы в прецентральной извилине как бы вверх ногами.
Рис.
4.1. Пирамидная система (схема).
А - Пирамидный путь: 1 - кора большого мозга; 2 - внутренняя капсула; 3 - ножка мозга; 4 - мост; 5 - перекрест пирамид; 6 - латеральный корково-спинномозговой (пирамидный) путь; 7 - спинной мозг; 8 - передний корково-спинномозговой путь; 9 - периферический нерв; III, VI, VII, IX, X, XI, XII - черепные нервы. Б - Конвекситальная поверхность коры большого мозга (поля 4 и 6); топографическая проекция двигательных функций: 1 - нога; 2 - туловище; 3 - рука; 4 - кисть; 5 - лицо. В - Горизонтальный срез через внутреннюю капсулу, расположение основных проводящих путей: 6 - зрительная и слуховая лучистость; 7 - височно-мостовые волокна и теменно-затылочно-мостовой пучок; 8 - таламические волокна; 9 - корково-спинномозговые волокна к нижней конечности; 10 - корково-спинномозговые
волокна к мышцам туловища; 11 - корково-спинномозговые волокна к верхней конечности; 12 - корково-ядерный путь; 13 - лобно-мостовой путь; 14 - корковоталамический путь; 15 - передняя ножка внутренней капсулы; 16 - колено внутренней капсулы; 17 - задняя ножка внутренней капсулы. Г - Передняя поверхность мозгового ствола: 18 - перекрест пирамид Аксоны мотонейронов формируют два нисходящих пути - корковоядерный,
направляющийся к ядрам черепных нервов, и более мощный - корковоспинномозговой, идущий к передним рогам спинного мозга. Волокна пирамидного пути, покидая двигательную зону коры, проходят через лучистый венец белого вещества мозга и конвергируют к внутренней капсуле. В соматотопическом порядке они проходят внутреннюю капсулу (в колене - корково-ядерный путь, в передних 2/3 заднего бедра - корково-спинномозговой путь) и идут в средней части ножек мозга, нисходят через каждую половину основания моста, будучи окруженными многочисленными нервными клетками ядер моста и волокнами различных систем.
На границе продолговатого и спинного мозга пирамидный путь становится видимым извне, его волокна образуют удлиненные пирамиды по обе стороны от средней линии продолговатого мозга (отсюда его название). В нижней части продолговатого мозга 80-85% волокон каждого пирамидного пути переходит на противоположную сторону, образуя латеральный пирамидный путь. Остальные волокна продолжают спускаться в гомолатеральных передних канатиках в составе переднего пирамидного пути. В шейном и грудном отделах спинного мозга его волокна соединяются с двигательными нейронами, обеспечивающими двустороннюю иннервацию мышц шеи, туловища, дыхательной мускулатуры, благодаря чему дыхание остается сохранным даже при грубом одностороннем поражении. Волокна, перешедшие на противоположную сторону, спускаются в составе латерального пирамидного пути в латеральных канатиках. Около 90% волокон образуют синапсы со вставочными нейронами, которые, в свою очередь, соединяются с большими α- и γ-мотонейронами переднего рога спинного мозга. Волокна, формирующие корково-ядерный путь, направляются к двигательным ядрам, расположенным в стволе мозга (V, VII, IX, X, XI, XII) черепных нервов, и обеспечивают двигательную иннервацию лицевой мускулатуры. Двигательные ядра черепных нервов являются гомологами передних рогов спинного мозга. Заслуживает внимания и другой пучок волокон, начинающийся в поле 8, обеспечивающем корковую иннервацию взора, а не в прецентральной извилине. Импульсы, идущие по этому пучку, обеспечивают содружественные движения глазных яблок в противоположную сторону. Волокна этого пучка на уровне лучистого венца присоединяются к пирамидному пути. Затем они проходят более вентрально в задней ножке внутренней капсулы, поворачивают каудально и идут к ядрам III, IV, VI черепных нервов.
Следует иметь в виду, что лишь часть волокон пирамидного пути составляет олигосинаптический двухнейронный путь. Значительная часть нисходящих волокон формирует полисинаптические пути, несущие информацию от различных отделов нервной системы. Наряду с афферентными волокнами, входящими в спинной мозг через задние корешки и несущими информацию от рецепторов, олиго- и полисинаптические модулируют активность двигательных нейронов (рис. 4.2, 4.3). Периферический мотонейрон. В передних рогах спинного мозга лежат мотонейроны - большие и малые а- и 7-клетки. Нейроны передних рогов мультиполярны. Их дендриты имеют множественные синаптические связи с различными афферентными и эфферентными системами.
Большие α-клетки с толстым и быстро проводящим аксоном осуществляют быстрые сокращения мышц и связаны с гигантскими клетками коры больших полушарий.

Малые а-клетки с более тонким аксоном выполняют тоническую функцию и получают информацию от экстрапирамидной системы. 7-Клетки с тонким и медленно проводящим аксоном иннервируют проприоцептивные мышечные веретена, регулируя их функциональное состояние. 7-Мотонейроны находятся под влиянием нисходящих пирамидных, ретикулярно-спинномозговых, вестибулоспинномозговых путей. Эфферентные влияния 7-волокон обеспечивают тонкую регуляцию произвольных движений и возможность регуляции силы ответа рецепторов на растяжение (система 7-мотонейрон - веретено).
Помимо непосредственно мотонейронов, в передних рогах спинного мозга имеется система вставочных нейронов, обеспечивающих
Рис.
4.2. Проводящие пути спинного мозга (схема).
1 - клиновидный пучок; 2 - тонкий пучок; 3 - задний спинно-мозжечковый путь; 4 - передний спинно-мозжечковый путь; 5 - латеральный спинно-таламический путь; 6 - спинно-покрышечный путь; 7 - спинно-оливный путь; 8 - передний спинноталамический путь; 9 - передние собственные пучки; 10 - передний корковоспинномозговой путь; 11 - покрышечно-спинномозговой путь; 12 - преддверноспинномозговой путь; 13 - оливо-спинномозговой путь; 14 - красноядерноспинномозговой путь; 15 - латеральный корковоспинномозговой путь; 16 - задние собственные пучки
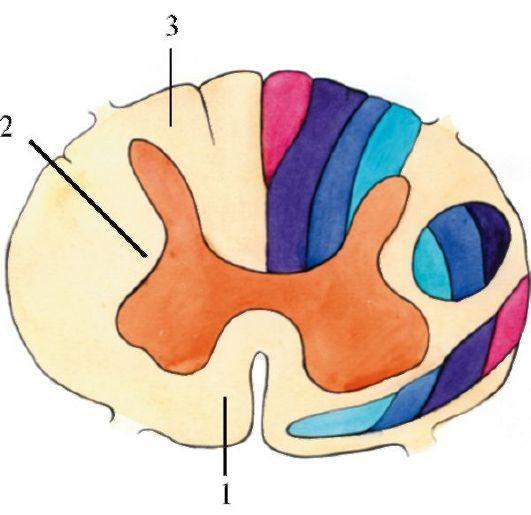
Рис.
4.3. Топография белого вещества спинного мозга (схема). 1 - передний канатик: синим цветом обозначены пути от шейных, грудных и поясничных сегментов, фиолетовым - от крестцовых; 2 - боковой канатик: голубым цветом обозначены пути от шейных сегментов, синим - от грудных, фиолетовым - от поясничных; 3 - задний канатик: голубым цветом обозначены пути от шейных сегментов, синим - от грудных, темно-синим - от поясничных, фиолетовым - от крестцовых регуляцию передачи сигналов от вышерасположенных отделов ЦНС,
периферических рецепторов, отвечающих за взаимодействие расположенных рядом сегментов спинного мозга. Некоторые из них оказывают облегчающее, другие - тормозящее действие (клетки Реншоу).
В передних рогах мотонейроны образуют группы, организованные в колонки в нескольких сегментах. В этих колонках имеется определенный соматотопический порядок (рис. 4.4). В шейном отделе латерально расположенные мотонейроны переднего рога иннервируют кисть и руку, а мотонейроны дистально лежащих колонок - мышцы шеи и грудной клетки. В поясничном отделе мотонейроны, иннервирующие стопу и ногу, также расположены латерально, а иннервирующие мускулатуру туловища - медиально.
Аксоны мотонейронов выходят из спинного мозга в составе передних корешков, объединяются с задними, формируя общий корешок, и в составе периферических нервов направляются к поперечнополосатой мускулатуре (рис. 4.5). Хорошо миелинизированные быстро проводящие аксоны больших а-клеток идут непосредственно к поперечнополосатой мускулатуре, формируя нервно-мышечные синапсы, или концевые пластинки. В состав нервов входят также эфферентные и афферентные волокна, исходящие из боковых рогов спинного мозга.

Волокно скелетной мышцы иннервируется аксоном только одного а-мотонейрона, но каждый а-мотонейрон может иннервировать разное число волокон скелетной мышцы. Количество мышечных волокон, иннервируемых одним α-мотонейроном, зависит от характера регуляции: так, у мышц с тонкой моторикой (например, глазные, артикулярные мышцы) один а-мотонейрон иннервирует лишь несколько волокон, а у
Рис.
4.4. Топография двигательных ядер в передних рогах спинного мозга на уровне шейного сегмента (схема). Слева - общее распределение клеток переднего рога; справа - ядра: 1 - заднемедиальное; 2 - переднемедиальное; 3 - переднее; 4 - центральное; 5 - переднелатеральное; 6 - заднелатеральное; 7 - зазаднелатеральное; I - гаммаэфферентные волокна от мелких клеток передних рогов к нервно-мышечным веретенам; II - соматические эфферентные волокна, дающие коллатерали к медиально расположенным клеткам Реншо; III - студенистое вещество

Рис.
4.5. Поперечный срез позвоночника и спинного мозга (схема). 1 - остистый отросток позвонка; 2 - синапс; 3 - кожный рецептор; 4 - афферентные (чувствительные) волокна; 5 - мышца; 6 - эфферентные (двигательные) волокна; 7 - тело позвонка; 8 - узел симпатического ствола; 9 - спинномозговой (чувствительный) узел; 10 - серое вещество спинного мозга; 11 - белое вещество спинного мозга мышц проксимальных отделов конечностей или у прямых мышц спины один α- мотонейрон иннервирует тысячи волокон.
α-Мотонейрон, его двигательный аксон и все иннервируемые им мышечные волокна образуют так называемую двигательную единицу, которая является основным элементом двигательного акта. В физиологических условиях разряд α-мотонейрона приводит к сокращению всех мышечных волокон двигательной единицы.
Скелетные мышечные волокна одной двигательной единицы называются мышечной единицей. Все волокна одной мышечной единицы принадлежат к одному и тому же гистохимическому типу: I, IIB или IIA. Двигательные единицы, сокращающиеся медленно и устойчивые к утомлению, классифицируются как медленные (S - slow) и состоят из волокон I типа. Мышечные единицы группы S обеспечиваются энергией за счет окислительного метаболизма, им свойственны слабые сокращения. Двигательные единицы, приводящие к быстрым фазическим одиночным сокращениям мышц, делятся на две
группы: быстрые утомляемые (FF - fastfatigable) и быстрые, устойчивые к утомлению (FR - fast fatigue resistant). Группа FF включает мышечные волокна типа IIB с гликолитическим энергетическим метаболизмом и сильными сокращениями, но быстрым утомлением. Группа FR включает мышечные волокна типа IIA с
окислительным метаболизмом и высокой устойчивостью к утомлению, сила их сокращения промежуточная.
Помимо больших и малых α-мотонейронов, передние рога содержат многочисленные 7-мотонейроны - клетки меньшего размера с диаметром сомы до 35 мкм. Дендриты γ-мотонейронов менее ветвисты и ориентированы преимущественно в поперечной плоскости. 7-Мотонейро- ны, проецирующиеся к конкретной мышце, расположены в том же двигательном ядре, что и α- мотонейроны. Тонкий медленно проводящий аксон γ-мотонейронов иннервирует интрафузальные мышечные волокна, составляющие проприорецепторы мышечного веретена.
Большие а-клетки связаны с гигантскими клетками коры полушарий большого мозга. Малые а-клетки имеют связь с экстрапирамидной системой. Через 7-клетки происходит регуляция состояния мышечных проприорецепторов. Среди различных мышечных рецепторов наиболее важными являются нервно-мышечные веретена. Афферентные волокна, называемые кольцеспиральными, или первичными, окончаниями, имеют довольно толстое миелиновое покрытие и относятся к быстро проводящим волокнам. Экстрафузальные волокна в расслабленном состоянии имеют постоянную длину. При растяжении мышцы растягивается веретено. Кольцеспиральные окончания реагируют на растяжение генерацией потенциала действия, который передается в большой мотонейрон по быстро проводящим афферентным волокнам, а затем опять по быстро проводящим толстым эфферентным волокнам - экстрафузальной мускулатуре. Мышца сокращается, ее исходная длина восстанавливается. Любое растяжение мышцы приводит в действие этот механизм. Постукивание по сухожилию мышцы вызывает ее растяжение. Немедленно реагируют веретена. Когда импульс достигает мотонейронов переднего рога спинного мозга, они реагируют, вызывая короткое сокращение. Эта моносинаптическая передача является базовой для всех проприоцептивных рефлексов. Рефлекторная дуга охватывает не более 1-2 сегментов спинного мозга, что имеет значение при определении локализации поражения.
Многие мышечные веретена имеют не только первичные, но и вторичные окончания. Эти окончания также отвечают на стимулы растяжения. Потенциал их действия распространяется в центральном направлении по тонким волокнам, сообщающимся со вставочными нейронами, ответственными за реципрокные действия соответствующих мышц-антагонистов.
Только небольшое число проприоцептивных импульсов достигает коры больших полушарий, большинство передается по кольцам обратной связи и не достигает коркового уровня. Это элементы рефлексов, которые служат основой произвольных и других движений, а также статических рефлексов, противодействующих силе тяжести.
Как при произвольном усилии, так и при рефлекторном движении в активность в первую очередь вступают наиболее тонкие аксоны. Их двигательные единицы генерируют очень слабые сокращения, что позволяет осуществлять тонкую регуляцию начальной фазы сокращения мышцы. По мере вовлечения двигательных единиц постепенно включаются α-мотонейроны с аксоном все большего диаметра, что сопровождается увеличением мышечного напряжения. Очередность вовлечения двигательных единиц соответствует порядку увеличения диаметра их аксона (принцип соразмерности).
Методика исследования Проводят осмотр, пальпацию и измерение объема мышц, определяют объем
активных и пассивных движений, мышечную силу, мышечный тонус, ритмику активных движений и рефлексы. Для установления характера и локализации
двигательных нарушений при клинически незначительно выраженных симптомах используют электрофизиологические методы.
Исследование двигательной функции начинают с осмотра мышц. Обращают внимание на атрофию или гипертрофию. Измеряя сантиметровой лентой окружность мышц, можно оценить выраженность трофических расстройств. Иногда можно заметить фибриллярные и фасцикулярные подергивания.
Активные движения проверяют последовательно во всех суставах (табл. 4.1) и выполняются обследуемым. Они могут отсутствовать или быть ограниченными в объеме и ослабленными. Полное отсутствие активных движений называют параличом, или плегией, ограничение объема движений или снижение их силы - парезом. Паралич или парез одной конечности носит название моноплегии, или монопареза. Паралич или парез обеих рук называют верхней параплегией, или парапарезом, паралич, или парапарез ног - нижней параплегией, или парапарезом. Паралич или парез двух одноименных конечностей, называют гемиплегией, или гемипарезом, паралич трех конечностей - триплегией, четырех конечностей - квадриплегией, или тетраплегией.
Таблица 4.1. Периферическая и сегментарная иннервация мышц