

35. Задачи экспериментальной полиплоидии, типы полиплоидов, используемые в селекции. Требования к объектам полиплоидии.
Полиплоидией называют изменчивость, связанную с кратным увеличением основного числа хромосом в клетках организма. Для полиплоидов характерно более мощное развитие вегетативных и репродуктивных органов, что вызвано увеличением у них размеров клеток. Полиплоидия играет большую роль в эволюции культурных растений. большинство родов покрытосеменных растений включают полиплоидные виды (1/3 от всех изученных видов). Ряд важнейших сельскохозяйственных культур представляют собой полиплоидные формы. Обычно более ценными оказываются формы с большим числом хромосом. Например, гексаплоидная мягкая пшеница, твёрдая пшеница (10–11% мировых посевов данной культуры). Диплоидная культурная однозернянка возделывается на незначительной площади. Полиплоидия в природе обнаружена у таких культур, как картофель (2n = 48), хлопчатник (2n = 26; 52), табак (2n = 48), арахис (2n = 20; 40), люцерна (2n = 16; 32; 48) и др. Однако у таких культур, как рожь (2n = 14), ячмень (2n = 14) и свёкла (2n = 18), в природе обнаружены только диплоидные виды.В селекции растений полиплоидию как практический приём стали применять после открытия А. Блексли в 1937 г. направленного полиплоидизирующего действия колхицина на делящиеся клетки растений. Колхицин – это алкалоид, выделенный из семян и клубнелуковиц безвременника осеннего.Экспериментальная полиплоидия позволяет решать следующие проблемы селекции и генетики растений:
1. Повышение продуктивности.2. Преодоление самонесовместимости.3. Преодоление межродовой и межвидовой нескрещиваемости.4. Восстановление плодовитости у отдалённых гибридов.5. Закрепление гетерозиса.6. Проведение синтеза и ресинтеза видов.7. Установление групп сцепления при генетическом анализе. В селекции используют два типа полиплоидов: аутополиплоиды и аллополиплоиды. Аутополиплоиды получают путём кратного увеличения в клетках наборов хромосом одного и того же вида (АА→АААА). Аллополиплоиды – соединением в одном геноме хромосомных наборов разных видов с последующим удвоением числа хромосом у гибридов F1 (АА х ВВ = F1 AB→ AABB). Аллополиплоиды получают также путём скрещивания разныхаутополиплоидов (АААА х ВВВВ = F1 ААВВ). Сегментные аллополиплоиды – это промежуточные между аутополи-плоидами и аллополиплоидами формы, у которых хромосомы разных геномов различаются не по всей длине, а частично.Основные различия между аутополиплоидами и аллополиплоидами: 1. фертильность наиболее сильно снижается у аутополиплоидов, что связано с нарушениями процесса мейоза, особенно у культур, возделываемых на семена. У аллополиплоидов хромосомы каждого типа представлены парами, поэтому мейоз протекает нормально и плодовитость снижается меньше, чем у аутополиплоидов.
2. Морфологически аутополиплоид сходен с родительской формой, а аллополиплоид занимает промежуточное положение между родительскими видами и похож на гибрид между ними. Известный шведский селекционер А. Леван (A.Levan, 1942) сформулировал основные положения, указывающие, какие растения наиболее перспективны для полиплоидизации.1. Растения с небольшим числом хромосом лучше реагируют на их удвоение, чем растения с высоким числом хромосом.2. Перекрёстноопыляющиеся культуры более подходят для селекции на основе полиплоидии, чем самоопылители. У них скорее восстанавливается фертильность.3. Чем более гетерозиготен материал, тем больше шансов получить выгодную новую комбинацию. Поэтому в качестве исходного материала для полиплоидизации целесообразно брать гибридный материал.4. Растения, у которых используются вегетативные части, представляют более перспективный материал для полиплоидии, чем выращиваемые на семена (Е.П. Раджабли, В.Д. Рудь, 1972).Кроме того, при использовании в селекции метода экспериментальной полиплоидии следует учитывать, что для разных видов растений характерен определённый оптимальный уровень плоидности (геномный оптимум), после достижения которого при дальнейшем увеличении числа хромосом интенсивность роста растений снижается. Например, удвоение числа хромосом у мягкой пшеницы приводит к слабому развитию растений. Наоборот, у ржи удвоение хромосом увеличивает мощность и продуктивность растений.
36. Положительные и отрицательные признаки полиплоидов, примеры культур, у которых используется аутополиплоидия и аллополиплоидия.К полезным изменениям при полиплоидии относят увеличение размеров органов и самих растений, повышение их продуктивности, содержания некоторых полезных веществ, устойчивости к полеганию и др.К нежелательным изменениям относится, прежде всего, снижение плодовитости аутополиплоидов. Они часто более позднеспелы, содержат больше воды в вегетативной массе. Характер наследования у аутополиплоидов сложнее, чем у диплоидов. Требуется больше времени для получения гомозиготного материала. С другой стороны, возможность поддержания селекционного материала полиплоидов в течение длительного времени в гетерозиготном состоянии можно использовать в селекции на гетерозис. При селекции полиплоидов важно иметь большой исходный материал, разнообразный в генетическом отношении. У самоопылителей целесообразно вовлекать в работу возможно большее количество линий при небольшом числе растений от каждой линии. У перекрёстников можно брать меньше семей, но с большим количеством растений в каждой семье. У вегетативно размножающихся растений и у тех растений, где бессемянность желательна, стерильность оказывается выгодной, поэтому в селекции этих культур полиплоидия оказывается перспективной. Например, полиплоидия используется при создании сортов нарцисса, тюльпана, гиацинта, хризантемы (тетраплоиды). Сорта винограда, чая, яблони являются также аутополиплоидами.Полиплоидия успешно используется для преодоления межвидовой несовместимости диких видов с культурными, например у картофеля.Использование аутополиплоидии в селекции культур, возделываемых на семена (рожь, гречиха) связано с определёнными трудностями, в частности со снижением плодовитости растений. При этом уменьшение общего количества семян на них обычно не компенсируется большей массой отдельного полиплоидного семени. Однако путём переопыления разных линий, их пересева и многократного отбора можно повысить фертильность тетраплоидов и увеличить их урожайность. Например, были получены устойчивые к полеганию, интенсивные сорта ржи: Стил (в Швеции), Белта, Ленинградская тетра и др. Сорт тетраплоидной ржи Тетра короткая (1986) создан путем удвоения числа хромосом у озимой ржи Короткостебельная 69, созданного путем ввода доминантного гена короткостебельности от сорта Болгарская низкостебельная в сорт Омка.В СибНИИСХе развивается направление селекции озимой тетраплоидной ржи, основанное на гибридизации тетраплоидных форм разного географического происхождения и последующих отборов. В качестве материнских форм используются сорта: Свеча, Белта, Пуховчанка, Крыжачек, Шатиловская тетра и др. В качестве отцовских форм (тестера) – Тетра гибрид, Тетра короткая, Популяция 2. Например, новый сорт тетраплоидной ржи Сибирь (1999) выведен в результате скрещивания трех тетраплоидных сортов: местная репродукция Белта, Тетра короткая, Шатиловская тетра.Тетраплоидные формы ржи создаются также на основе диплоидных гибридов первого поколения с целью закрепления гетерозисного эффекта переводом гибридов на полиплоидный уровень. Наиболее доступный и эффективный метод удвоения числа хромосом – обработка семян, проростков или колосьев раствором колхицина (Р.И. Рутц, 1990). Созданы также тетраплоидные сорта гречихи с дружным созреванием, хорошей озернённостью, с более низким стеблестоем, устойчивые к полеганию и осыпанию, с повышенным содержанием белка в зерне (13–16%): Искра, Минчанка, Большевик 4 и др.Переопыление полиплоидных сортов с диплоидными приводит к резкому снижению семенной продуктивности (особенно у тетраплоидов). Поэтому должна быть пространственная изоляция между ди-плоидными и полиплоидными сортами одной культуры.У растений, размножающихся семенами, но возделываемых для получения их вегетативных органов, пониженная плодовитость не имеет решающего значения. Например, тетраплоидные сорта ряда кормовых и овощных культур: турнепс, свёкла, редис, укроп, шпинат. Положительными свойствами обладают также аутотетра-плоиды люцерны, эспарцета, тыквы и других культур. Использование аллополиплоидов. Ценность аллополиплоидов не находится в прямой связи с хозяйственными достоинствами объединённых в них видов. О свойствах аллополиплоида можно судить по качеству обычного диплоидного гибрида, возникшего от скрещивания тех же родительских форм. Наиболее удачны те аллополиплоиды, которые получают от полностью стерильных гибридов F1. К природнымаллополиплоидам, размножающимся семенами, относят большинство видов пшеницы, овса, хлопчатника, землянику обыкновенную, сливу и др. Практическая ценность экспериментальных аллополиплоидов зависит от цитоплазмы, на основе которой они создаются, а также от удачного выбора биотипов родительских видов, обеспечивающих наилучшую комбинацию признаков.
37.Факторы, вызывающие полиплоидию, колхициновые методы получения полиплоидов.Для искусственного индуцирования полиплоидии используются различные факторы, которые в основном вызывают полиплоидизацию и в природе. Физические – температурные воздействия, ионизирующие излучения. Механические – повреждение тканей, центрифугирование. Из химических веществ применяют колхицин, аппиоль (экстракт из семян петрушки), ауранцию (в 100 раз эффективнее колхицина), закись азота (этот газ применяют под давлением в несколько атмосфер, воздействуя на цветки после оплодотворения). Для получения полиплоидов используют также аценафтен, хлористыйсангуинарин, гаммексан, линдан и другие вещества. Из ранних приёмов экспериментального получения полиплоидов наиболее эффективными были методы декапитации и воздействия высокими температурами. Сущность метода декапитации сводится к возможности получения тетраплоидных побегов из регенерирующей ткани каллюса после удаления верхушки растения. Этим методом получены тетраплоиды томатов, капусты и других культур. Под воздействием высокой температуры на зиготу в стадии первого деления были получены тетраплоидные формы у кукурузы, ржи, пшеницы, ячменя, донника белого, люцерны и льна. Полиплоиды можно выделить из близнецовых зародышей, возникающих в результате естественной полиэмбрионии. Новые методы получения полиплоидов основаны на использовании генетического контроля над процессами полиплоидизации. Например, у кукурузы известен рецессивный ген elongate, который в гомозиготном состоянии вызывает образование диплоидных яйцеклеток. Этот ген можно ввести в любую инцухт-линию. Для индуцирования полиплоидии в процессе оплодотворения используют также гены асинапсиса. Действие этих генов приводит к образованию нередуцированных гамет и как следствие – к возникновению полиплоидов.К генетическим методам относится получение полиплоидов в результате межвидовых и межродовых скрещиваний, которые приводят к аномальному прохождению мейоза у гибридов.В настоящее время в селекции для получения полиплоидов чаще используют колхицин в виде водных растворов, ланолиновой пасты и в виде раствора в агаре или глицерине. В чистом виде колхицин представляет собой желтовато-белый порошок, растворимый в воде, спирте и хлороформе. Химическая формула колхицина – С22Н25О6N.Получение полиплоидов с помощью алкалоида колхицина основано на том, что это соединение оказывает наркотическое действие на делящиеся клетки, препятствуя расхождению в митозе сестринских хромосом к противоположным полюсам и образованию дочерних клеток. В результате клеточная перегородка не образуется и сама клетка не делится. Удвоенные хромосомы остаются в одной исходной клетке. Когда наркотическое действие проходит, клетка делится и даёт начало двум тетраплоидным клеткам. Таким образом, действие колхицина заключается в подавлении функции веретена деления, в результате чего парализуется расхождение дочерних хромосом к полюсам и они остаются в одной клетке.Обычно готовят 1–2%-ный раствор, а затем его разбавляют до нужной концентрации. Колхицин характеризуется большой стойкостью, поэтому его растворы можно стерилизовать в автоклаве. Маточный раствор хранят в темноте, т. к. на свету колхицин разлагается. При работе соблюдают осторожность, т. к. колхицин – сильный яд. С учётом этих условий разработан ряд методов получения полиплоидов для разных культур.1.Колхицинирование семян. Этот способ пригоден для культур с быстропрорастающими семенами. Семена подвергают обработке либо в сухом виде, либо предварительно замачивают в воде. Перед посевом семена промывают в проточной воде. Концентрация раствора 0,1–0,2%, экспозиция – 3–6 дней. Если предварительно замачивают в воде, то набухшие семена проращивают в чашках Петри в течение 0,5–48 ч на фильтровальной бумаге, смоченной колхицином.Например, таким путём получают тетраплоиды картофеля, табака, райграса, клевера, шелковицы и других культур. Положительная сторона метода колхицинирования семян – почти полное отсутствие химерных тканей у выросших растений. Недостаток – резко снижается выживаемость проростков вследствие задержки развития корневой системы. Это не желательно при ограниченном количестве семян (при отдалённой гибридизации).2.Погружение проростковв водный раствор колхицина или помещение их на фильтровальную бумагу, смоченную колхицином. Концентрация раствора 0,01–0,2%, продолжительность обработки 3–12 ч и более. Недостаток – сильная задержка в развитии и гибель проростков (семян). Для устранения этого недостатка корешки изолируют от действия раствора колхицина. Для этого проросшие семена укрепляют на специальной сетке корешками вверх. Например, у зерновых злаков проростки с колеоптилем длиной 2–4 мм опускают на 30 мин в чашку Петри корешками вверх. После этого их высаживают в ящики в теплице. 3.Обработка мелкосемянных культур.семена проращивают в чашках Петри на фильтровальной бумаге. В момент наклёвывания семян чашки переворачивают вверх дном и отрастающие корешки в результате геотропизма растут вниз. Когда они достигнут 0,5–0,8 см длины, чашки возвращают в исходное положение, семена заливают раствором колхицина, а корешки накрывают влажной фильтровальной бумагой. Концентрация раствора 0,05–0,1%, экспозиция – 2 ч. Этим методом получают тетраплоиды моркови, салата, петрушки и других культур.4.Капельный метод является одним из самых надёжных для двудольных растений. Суть его в том, что капли раствора колхицина наносят пипеткой на точку роста молодых сеянцев утром и вечером или через каждые 3–4 ч в течение 3–4 сут, иногда с перерывом на несколько суток. При этом используют водные растворы колхицина, водно-глицериновые и водно-агаровые (0,4% агара). Концентрация раствора 0,1–0,4%. Растения при обработке содержат на рассеянном свету, относительная влажность воздуха 70–80%. Лучше вместо водных растворов использовать колхицин-трагакантовую смесь, т. к. вода быстро испаряется. Эта смесь хорошо прилипает, не смывается при поливе.5.Метод инъекций. При работе со злаковыми культурами раствор колхицина концентрацией 0,1–0,2% вводят шприцем в центральную часть стебелька на уровне корневой шейки. У кукурузы обработку этим способом проводят в фазе 1–2 листа в утренние часы. Впрыскивание заканчивают, когда вверху, в раструбе развивающегося листа, появляется капля раствора. Обработку повторяют в течение нескольких дней. Для пшеницы разработан метод полиплоидизации путём инъекции 0,1–1,0%-ного раствора колхицина в цветки на трёх стадиях развития: до опыления, во время опыления и после опыления. У винограда инъекции проводят в молодые побеги, у капусты – в почки маточных растений на ранних стадиях их развития.6. Метод CIMMYT. В CIMMYT при получении тритикале используют метод введения раствора колхицина в растение с помощью глазных пипеток с укороченным концом через основания листочков. 7.Обработка корней. Этот метод наиболее эффективен при работе с пшеницей, просом и другими злаками, у которых верхушка малодоступна, а также при работе с гречихой, томатами и прочими культурами. В начале выкапывают молодые растения и отмывают корни, затем попеременно погружают на 12 ч то в слабый раствор колхицина, то в проточную воду для снижения повреждения корней. Концентрация раствора – 0,0125–0,4%, экспозиция – 24–144 ч.8.Обработка взрослых растений. Суть метода заключается в том, что оставляют несколько побегов, на которых обрабатывают все точки роста. Концентрация раствора выше обычного (0,2–1%). Обработку можно проводить, нагибая побеги и погружая их в раствор. При этом используют капельный метод, тампоны и желатиновые капсулы, метод инъекций и др.9.Обработка путем погруженияпобега. На побеге делают небольшой надрез на 1–2 см ниже верхушки и погружают надрезанную часть в пробирку с раствором. Все почки на расстоянии не менее 4–5 см от обработанной части удаляют. 10.Обработка цветоносных побегов. Суть метода в том, что колхицин вводят в растение через стебель в период заложения и формирования спорогенной ткани. Применяется метод у двулетних культур (сахарная и кормовая свёкла, турнепс). Обработка цветоносных побегов во второй год жизни ускоряет работу, т. к. позволяет получить тетраплоидные семена в первый год жизни. Например, у свёклы надрезают до половины у основания цветоносный побег (длиной 10–12 см) и расщепляют его. Отщепленный конец погружают в пробирку с 0,01%-ным раствором колхицина. В результате образуются диплоидные яйцеклетки и пыльца. Таким путём удаётся получить до 40–50% тетраплоидных семян.
38.Создание триплоидных гибридов сахарной свёклы.Триплоидные гибриды сахарной свёклы стали создавать после того, как Ф. Пето и Дж. Бойс показали, что для триплоидов характерно большое содержание сахара в соке, большая масса корня и высокий выход сахара на единицу площади посева по сравнению с диплоидами. Для получения гибридных семян скрещивают диплоидные и тетраплоидные формы. Семенники диплоидов и тетраплоидов высаживают чередующимися рядами, и в результате естественногопереопыления образуются гибридные триплоидные семена. Наиболее рентабельно размещение диплоидных и тетраплоидных форм в отношении 1 : 3. Поскольку пыльцевые трубки диплоидных форм растут быстрее, чем тетраплоидных, то наблюдается тенденция к оплодотворению пыльцой диплоидных растений, поэтому на тетраплоидных растениях будут преобладать триплоидные семена. При уборке одновременно со всего участка без вычленения родительских компонентов получают около 70–75% триплоидных семян в ворохе. При раздельной уборке с каждой из родительских форм доля триплоидных семян на тетраплоидных растениях может достигать 90%. Например, районированный в 1987 г. в Хмельницкой области гибрид Белоцерковский поли 41 получают скрещиванием односемянной тетраплоидной формы свёклы с диплоидной многосемянной при соотношении компонентов 3 : 1. Уборку проводят раздельно и используют семена только односемянного компонента.Повысить выход триплоидных семян можно, если в качестве материнского компонента использовать растения с ЦМС. Триплоидные гибриды сахарной свёклы дают с единицы площади на 15–20% больше сахара, чем диплоидные сорта, характеризуются устойчивостью к церкоспорозу. У этих гибридов ослаблена отрицательная корреляция между увеличением размеров корнеплодов и повышением их сахаристости. В нашей стране селекция триплоидных гибридов началась в 1958 г. в ИЦиГе (Новосибирск). Первые два отечественных триплоидных гибрида сахарной свёклы – Кубанский полигибрид 9 и Белоцерковский полигибрид 1 были районированы в 1964 г.
Селекция одноростковой сахарной свёклы начата в 1932 г., когда О.К. Коломиец во ВНИИСе в результате обследования семенников сорта Верхнячская 1025 обнаружил куст с наличием односемянных плодов. В 1966 г. районирован первый гибрид одноростковой свёклы Белоцерковский полигибрид 2, превзошедший стандарт по сбору сахара с гектара и урожаю корней, он даёт 70–75% одноростковых семян.Возделывание одноростковых гибридов сахарной свёклы позволяет применять механизированную прорывку всходов, что значительно снижает затраты труда. Обнаружение Ф. Оуэном в 1942 г. мужской стерильности у свёклы открыло возможность для более полного использования преимуществ триплоидов. Для производства триплоидных семян сахарной свёклы на стерильной основе необходимо иметь следующие компоненты:1. Диплоидный сорт с ЦМС – 2n (2x) ЦМС, (S) XXZZ. 2. Закрепитель мужской стерильности – О-тип 2n (2x), (N) xxzz.3. Тетраплоидный опылитель – 2n (4x), XXZZ. Большой практический интерес представляют гетерозисные триплоидные гибриды от скрещивания тетраплоидной кормовой свёклы с диплоидной сахарной или тетраплоидной сахарной с диплоидной кормовой свёклой. При этом выход сахара с единицы площади может достигать рекордных показателей. Например, сахарно-кормовой гибрид Тимирязевский 12. Исследования показали, что высокоурожайные сорта и гибриды сахарной свёклы могут быть получены и на тетраплоидном уровне. Тетраплоиды могут давать равнозначный с триплоидами гетерозисный эффект. Это даёт возможность использовать у тетраплоидных гибридов продлённый гетерозис. Данный эффект обнаружен также у кормовой свёклы и её гибридов с сахарной свёклой.На том же принципе, что и получение триплоидных гибридов свёклы, основан метод производства бессемянных триплоидных арбузов (2n = 33), разработанный японским генетиком Х. Кихарой в 1951 г. Их получают путём скрещивания диплоидных (2n = 22) и тетраплоидных (2n = 44) форм арбуза. У такого гибрида полностью нарушен мейоз и триплоидные растения не образуют семян, что удобно для использования в пищу.
39. Признаки полиплоидизации растений, ход селекционного процесса с использованием полиплоидии. Получение тетраплоидных сортов ржи и гречихи. Выделенные в год обработки полиплоидные растения обозначают как С0, а их потомства – С1, С2 и т. д. Работу проводят в специальном питомнике. У перекрёстноопыляющихся культур чаще получают тетраплоиды. При выбраковке не следует удалять целое растение, необходимо анализировать каждый побег, особенно у двулетних растений, клевера и злаков. Это связано с тем, что чаще всего полиплоидные растения являются химерами. Диплоидные побеги растут быстрее и их сразу выбраковывают. Тетраплоидные цветоносные побеги более позднеспелые и позже зацветают. Например, у ячменя тетраплоидные побеги из узла кущения появляются позже диплоидных. В поколении C0 для растений должны быть созданы благоприятные условия выращивания: высокая влажность воздуха, оптимальная температура, не очень сильное освещение на ранних этапах (затеняют всходы). Для клевера и злаков применяют разреженный посев, для того чтобы вызвать максимальное кущение, которое приводит к большему появлению тетраплоидов.Как правило, для выделения тетраплоидов в C0 исследователи применяют косвенные методы определения плоидности. Полиплоидные растения обнаруживают и выделяют по ряду признаков. Листья у полиплоидов крупнее, шире и толще. Полиплоиды отличаются крупными тёмно-зелёными семядолями с укороченным подсемядольным коленом. У однодольныхколеоптиль утолщён и укорочен, первые листовые пластинки тёмно-зелёные, утолщённые и гофрированные с нарушенным жилкованием. Используют также признаки, относящиеся к репродуктивным органам: размер цветков, бутонов, колосьев, пыльников.Стабильным показателем, не зависящим от условий выращивания, является размер пыльцевых зёрен и число микропор наихэкзине. Отбор тетраплоидов по размеру пыльцы с успехом применяется у свёклы, капусты, турнепса, редиса, редьки, ржи, гречихи, клевера и других культур. Отбор тетраплоидов по числу микропор на экзине пыльцы проводят при помещении пыльцевых зёрен в концентрированную серную кислоту. У маревых и капустных гаплоидная пыльца содержит на экзине 3 микропоры, диплоидная – 3–4. Для злаков данный показатель не пригоден, т. к. у пыльцевых зёрен любой плоидности только одна микропора.В первом семенном поколении возможен отбор семей полиплоидов по размерам семян. Например, масса 1000 семян у диплоидной ржи – 29,6 г, а у тетраплоидной – 46,3 г; у проса соответственно 5,2 и 8,6 г; у клевера – 1,8 и 3,4 г. Эти различия позволяют отделить полиплоидные семена путём фракционирования на решетах, например, у клевера, редиса и капусты.Отбор до цветения можно проводить по анатомическим показателям – клеткам эпидермиса, размеру и числу устьиц на единицу листовой поверхности, числу хлоропластов в замыкающих клетках устьиц. У тетраплоидов замыкающие клетки устьиц гораздо крупнее, число хлоропластов в них в 1,5–2 раза больше, чем у диплоидов, и нередко они расположены в два ряда. Более точным показателем полиплоидного состояния является число хлоропластов в замыкающих клетках устьиц. Поскольку из-за химерности листья и цветки могут иметь разную плоидность в C0, то отбор по этим признакам можно проводить только в C1, когда химерность отсутствует. Для этого отдельно высевают потомство каждого растения C0. После появления всходов растения маркируют, берут отрезки листьев, фиксируют в уксусномалкаголе, окрашивают 5%-нымJ в KJ и просматривают в капле 45%-ной уксусной кислоты под микроскопом. Для анализа лучше брать жёлтые листья, т. к. они не перекрашиваются. Наиболее надёжный метод определения плоидности – микроскопическое исследование клеток корешков или конуса нарастания развивающихся побегов. Цитологический анализ лучше проводить с поколения C1, проращивая часть семян из каждой семьи.Большая часть искусственно полученныхполиплоидов не имеет непосредственного хозяйственного значения. Некоторые первичные «сырые» полиплоиды имеют прямую практическую ценность. В большинстве случаев новые формы нуждаются в улучшении. Это достигается путём генетической рекомбинации при скрещивании их между собой или путём отбора, особенно у самоопылителей. У аллогамных видов улучшение может происходить естественным путём.
40. Роль анеуплоидов в селекции. Методы получения гаплоидов и пути использования гаплоидии в селекции.Анеуплоиды, в частности нуллисомики (2n – 2), моносомики (2n – 1), трисомики (2n + 1) и другие, в настоящее время применяют в генетическом анализе, результаты которого используют в практической селекции. Например, у мягкой пшеницы после создания серий моносом-ных и трисомных линий стало возможно определять генный состав её хромосом, а также локализовать любой ген и осуществить замещение одних хромосом другими. Это было проведено Э. Сирсом у сорта ЧайнизСпринг (21 моносомная линия). В результате этих работ га-плоидный набор мягкой пшеницы был разбит на семь гомеологических групп, по три хромосомы в каждой группе в зависимости от их принадлежности к геномам А, В и D. Полные ряды моносомиков и трисомиков созданы у пшеницы, овса и некоторых других культур. Серии моносомных и трисомных линий используются для изучения наследования различных признаков с учётом генетического вклада каждой отдельной хромосомы. Благодаря таким исследованиям открывается возможность манипулировать с отдельными хромосомами или их фрагментами (генетическая инженерия). Например, сорту яровой пшеницы Тетчер, у которого в хромосомах III, XIII, XIX содержится по одному гену устойчивости к бурой ржавчине, путём замещения хромосом X, XX удалось добавить ещё два гена резистентности. У диплоидных культур создают серии трисомиков. Гаплоиды – это особи обычно диплоидных или аллополиплоидных видов, в соматических клетках которых содержится в два раза меньше хромосом, чем у исходных форм, причём из каждой пары гомологичных хромосом представлена только одна хромосома. Гаплоидия применяется для изучения генетики и эволюции растений. С её помощью определяют геномный состав видов и уточняют их таксономическое положение, исследуют влияние дозы геномов в полиплоидных рядах, выясняют происхождение и генетические причины апомиксиса. На явлении гаплоидии основаны методы получения из гаплоидов гомозиготных диплоидных линий, а также удачных рекомбинаций генов при комбинационной селекции. Впервые на перспективность применения гаплоидов для решения селекционных задач указал в 1929 г. Г.Д. Карпеченко. Поскольку в гаметах желательное сочетание генов бывает чаще, чем в зиготах, то проще и быстрее получить гомозиготную форму с нужным сочетанием признаков путём прямого удвоения хромосом у гаплоидов. Например, для получения всех возможных типов гомозиготных линий на основе перекомбинации семи пар независимых генов необходимо перевести на диплоидный уровень только 128 гамет гибридов F1 (27). На диплоидном уровне для достижения такого же результата потребуется уже не менее 16384 рас-тенийF2 (47). То есть эффективность комбинационной селекции во много раз возрастает при разработке способа массового выделения гаплоидов для последующего перевода их на диплоидный уровень. Путём удвоения числа хромосом у гаплоидов можно сразу создать гомозиготные линии (у перекрёстноопыляющихся культур на выделение их требуется до 7–10 лет).
Для экспериментального получения гаплоидных растений также используют следующие методы: 1) культивирование пыльников и микроспор в стерильных условиях на питательной среде; 2) отбор близнецов; 3) межвидовые скрещивания; 4) радиологический метод; 5) гибридизация на разных уровнях плоидности.Наиболее перспективными методами являются культивирование пыльников и пыльцы (микроспор) в стерильных условиях (invitro).С. Гуха и С. Махашвари в 1964 г. впервые обнаружили в культуре invitro пыльников дурмана (Datura) эмбриоподобные структуры. Через два года из такой структуры было получено гаплоидное растение. Авторы указали на происхождение этого растения из гаплоидного пыльцевого зерна. Большой вклад в разработку метода массового получения гаплоидов внёс Дж. П. Нич с сотрудниками. Разработанный ими в 1972 г. метод лёг в основу использования культуры пыльников для вызывания invitroандрогенеза у растений. Г. Мельхерс (1976) наметил новые пути массового получения гаплоидов в селекции растений и внёс предложение соединить метод культуры пыльников с общепринятыми методами селекции.Для получения дигаплоидных растений в процессе селекции вначале проводят скрещивания. Из пыльников растений F1 получают гаплоиды, удваивают у них число хромосом с помощью колхицина. Потомства полученных дигаплоидных растений оценивают в поле в F3 и F4, а лучшие линии проходят испытания на продуктивность в F5–F8 (рис. 7.1).При культуре пыльников редко удаётся полностью исключить размножение диплоидных клеток ткани пыльника и возникновение из них диплоидных эмбрионов. Поэтому более перспективно получение гаплоидов из изолированной пыльцы.
41. Понятие о селекции на гетерозис, типы гибридов кукурузы, используемые в производстве. Пути снижения стоимости гибридных семян Гетерозис в широком смысле – это все положительные эффекты, ведущие к превосходству гибридов F1 над родительскими формами. Гетерозис в полной мере проявляется в F1. При генеративном размножении в последующих поколениях он теряется. У вегетативно размножаемых растений гетерозис передаётся потомству стойко. Для практического использования гетерозиса у генеративно размножаемых видов необходимо в больших масштабах скрещивать определённые родительские формы (линии, сорта). Селекцией на гетерозис называют создание гибридов F1, отличающихся высоким гетерозисом по урожайности, качеству продукции и другим хозяйственно важным признакам. При селекции на гетерозис скрещивание служит для массового получения гибридных семян и их дальнейшего использования в производстве. Гетерозисные гибриды по урожайности превышают обычные свободноопыляющиеся сорта на 30–50%. Явление гетерозиса широко используют в селекции кукурузы, сорго, подсолнечника, томата и других культур. Явление гетерозиса объясняется тремя основными гипотезами:1. Гипотеза сверхдоминирования объясняет проявление гетерозиса гетерозиготным состоянием гибридов.2. Гипотеза доминирования исходит из того, что к гетерозису ведёт вызванное скрещиванием накопление доминантных аллелей продуктивности.3. Гипотеза генетического баланса объясняет гетерозис физиологической сбалансированностью процессов обмена веществ. Различают следующие типы гибридов производственного использования:1. Межлинейные (простые – скрещивают две самоопылённые линии; трёхлинейные – простой межлинейный гибрид х самоопылённую линию; двойные – скрещивают два простых межлинейных гибрида; сложные межлинейные – скрещивают более четырех самоопылённых линий).2. Сортолинейные (простые – сорт х линию; сложные – сорт х простой межлинейный гибрид).3. Линейно-сортовые – простой гибрид х сорт.4. Межсортовые гибриды.5. Гибридные (синтетические) популяции получают путём смешения семян простых гибридов и их свободного переопыления. Наиболее высокую прибавку урожая дают гибриды, полученные с участием самоопылённых линий. Поэтому в производстве распространены в основном межлинейные и сортолинейные гибриды кукурузы и других культур.У аутогамных (самоопыляющихся) видов – пшеницы, риса, ячменя и других, преобладающей формой гетерозисной селекции является создание межсортовых гибридов.У кукурузы лучшие межлинейные гибриды по продуктивности превышают свободноопыляемые сорта на 30–40%. Эти гибриды отличаются высокой гомогенностью, но они очень дорогостоящие.Снизить высокую стоимость производства простых межлинейных гибридов можно несколькими способами:1. удачный выбор исходного материала может уменьшить инцухт-депрессию линий и повысить их урожайность.2. Использование сестринских скрещиваний (А х А1) х (В х В1), где А1 и В1 – сестринские линии.3. скрещивание двух простых гибридов (А х В) х (С х Д).Двойные гибриды имеют почти такую же урожайность, как и простые, но уступают им по выравненности. Они характеризуются лучшей приспособленностью к окружающей среде.
42. Методы создания самоопылённых линий.Гетерозисная селекция включает несколько этапов:1. Подбор исходного материала.2. Создание самоопылённых линий.3. Изучение комбинационной способности этих линий.4. Использование наиболее пригодных линий в качестве родительских форм для создания простых, двойных и других типов гибридов или для составления гибридной популяции.Исходный материал подбирают путём диаллельного скрещивания ряда неродственных сортов или популяций. Сорта или популяции, которые дают наивысший эффект гетерозиса, используются для заложения питомника самоопылённых линий. Из популяций исходного материала с хорошей комбинационной способностью можно получить самоопылённые линии с отличной комбинационной способностью. Испытание исходного материала на комбинационную способность повышает затраты труда и времени, и поэтому используют косвенные критерии для подбора исходного материала.
1. Высокой комбинационной способности можно добиться, когда самоопылённые линии получают из использования популяций разного географического происхождения. (Разное географическое происхождение популяций, из которых получают самоопыленные линии).2. скрещивание разных видов или разновидностей (гибриды кремнистых и зубовидных форм кукурузы с эффектом гетерозиса 50%).3. Использование линий, полученных из межлинейных и сортолинейных гибридов. Однако при создании раннеспелых гибридов рекомендуют отбирать линии из сортов или межсортовых гибридов.При использовании данных критериев можно добиться комбинационной способности скрещиваемых форм.Применяется ряд методов получения самоопылённых линий. При использовании любого из них необходимо иметь питомник отбора, включающий до 500–1000 сортов или гибридов, где производят самоопыление. Самоопылению подвергают не менее 250 растений.1. Стандартный метод – это разновидность индивидуального отбора, при котором отбираются лучшие по комплексу признаков и свойств растения, потомства которых самоопыляются и оцениваются до тех пор, пока не будут получены однородные гомозиготные линии. Схема метода следующая:1-й год – питомник отбора.
2-й год – отбор лучших семян I1, посев и самоопыление лучших растений I1 (худшие потомства бракуются).3-й год – браковка худших семей I2, отбор и самоопыление лучших растений в потомствах.4-й год – одну часть семян I3c каждого растения высевают в питомнике линий, где продолжаются их самоопыление и оценка, а вторую часть семян используют для определения общей комбинационной способности (ОКС) линий.5-й год – проводят предварительное испытание и отбор наиболее ценных линий и изучают гибриды от скрещивания линий с тестером.6-й год – отбирают лучшие линии и самоопыляют растения, а также продолжают испытание гибридов.7-й год – на основе ОКС отбирают лучшие линии I6, которые включают в диаллельные скрещивания для получения высокогетерозисных гибридов.2. Гнездовой метод – это вариант стандартного метода, заключающийся в том, что семена от каждого самоопылённого початка на следующий год высеваются по 3–4 шт. в гнездо для оценки. (Метод предложен Джонсом и Синглетоном в 1934 г.) Проводится отбор растений, отвечающих модели создаваемой линии. Этот метод особенно эффективен в том случае, если комбинационную способность линий определяют, начиная с I2. Данный метод позволяет испытать на комбинационную способность в 10–20 раз больше семей и линий по сравнению со стандартным методом при одних и тех же затратах труда.3. Рекуррентный (периодический) отбор – это отбор в популяциях перекрёстноопыляющихся растений, обеспечивающий последовательное повышение концентрации нужных комплексов наследственных факторов. Данный метод предусматривает периодическое чередование приёмов выделения лучших генотипов путём инцухта и скрещивания их для получения рекомбинаций. Процедура периодического отбора состоит в следующем. В первый год отобранные в исходном гибриде или сложной популяции 150–200 лучших растений скрещивают с тестером, т.е. с лучшей самоопылённой линией. Параллельно размножают эти растения при самоопылении. Во второй год проводят сравнительное испытание гибридов от скрещивания отобранных растений с тестером. По данным испытания отбирают 5–10% лучших гибридов. На третий год самоопылённые семена 5–10% лучших гибридов высевают отдельными рядками и проводят скрещивания во всех возможных комбинациях. Гибридные семена объединяют и выращивают совместно. Далее на этом посеве отбирают растения для следующего цикла, при этом используют тот же тестер.Метод рекуррентного отбора широко применяют, например, в селекции кукурузы для получения линий с высокой специфической комбинационной способностью.Преимущество рекуррентного отбора в том, что уже с первого цикла отбора важнейшим критерием служит комбинационная способность. Однако затраты средств и времени выше, чем при использовании стандартного метода.4. Метод гаплоидии – получение форм растений с половинным набором хромосом в клетках – гаплоидов. Путём удвоения хромосом у гаплоидов получают гомозиготные линии, что позволяет резко сократить время, затрачиваемое на создание линий.Для улучшения существующих линий по разным признакам применяют методы комбинационной и мутационной селекции. Беккроссы (возвратные скрещивания) применяют для передачи генов-восстанови-телей фертильности и генов устойчивости к болезням, при переводе линий на ЦМС. Если нужно ввести в линии количественный признак, то используют метод педигри. Мутагенные факторы позволяют вызвать у растений изменение отдельных признаков, которые можно использовать в гибридизации (короткостебельность, устойчивость к болезням, улучшенные качества и др.). Для улучшения самоопылённых линий применяют также метод отбора гамет, метод конвергентной селекции и др.
43. Схемы создания двойных гетерозисных гибридов на основе использования ЦМС. Типы ЦМС и самоопылённых линий. Примеры трёхлинейных гибридов кукурузы.Не каждое скрещивание двух линий приводит к получению гетерозисных гибридов. Чтобы найти наиболее пригодные для этого линии, необходимо проводить диаллельные скрещивания. Для сокращения объёма работы разбивают все линии на небольшие группы (8–10 линий) и диаллельные скрещивания проводят в пределах каждой из этих групп в отдельности. Полученные гибриды испытывают в полевом опыте по урожайности и другим признакам. Оценка на комбинационную способность линий на основе испытания из гибридов F1 может быть проведена по двум критериям:1. По ОКС, с помощью которой выявляют линии, в среднем давшие наиболее урожайные гибриды. ОКС относится к одной конкретной линии.2. По СКС, которая показывает, какие комбинации двух линий дают гибриды F1c наивысшей урожайностью, достоверно превышающей тот уровень урожайности, который следовало ожидать на основе ОКС двух данных родительских форм (линий). Следовательно, СКС относится к паре линий, а не к отдельно взятым линиям. На первых этапах получения линий их оценку проводят только на ОКС, так как СКС определяют с помощью трудоёмкого диаллельного скрещивания.
ОКС определяют с помощью топкроссных испытаний. Топкроссом называют скрещивание некоторого числа линий с определённым сортом-тестером. В качестве сорта-тестера используют свободноопыляемую (у перекрёстников) популяцию с широкой генетической основой. Сорт-тестер по возможности должен обладать средней ОКС. Для повышения надёжности определения ОКС топкроссные испытания проводят с несколькими сортами-тестерами.
ОКС экологически более стабильна, чем СКС. Для определения ОКС высевают по одному рядку каждой испытываемой линии. Каждые 2–3 рядка линий чередуют с рядом сорта-тестера. Как правило, тестером служит отцовская родительская форма. Кастрированные материнские растения опыляют пыльцой тестера. При использовании тестера в качестве материнской формы все гибриды имеют одинаковые по размеру семена.
После создания генотипически стабильных самоопылённых линий в поколении I5 или I6 необходимо провести испытание лучших линий на СКС методом диаллельного анализа. В этом испытании участвуют наиболее урожайные комбинации линий. При определении СКС гибриды F1 испытывают в разных местах и в разные годы или сезоны, поскольку СКС больше, чем ОКС подвержена влиянию взаимодействий генотипа со средой.
После выявления пар линий с высокой СКС на основе хорошей ОКС селекционный процесс заканчивается, если цель селекции – создание простых гибридов.Подбор комбинаций двойных гибридов. По экономическим причинам во многих странах в производстве ещё преобладают двойные гибриды, поэтому выявляют простые гибриды, наиболее пригодные для создания двойных межлинейных гибридов. Для получения высокоурожайного и экологически пластичного двойного гибрида необходимо подвергнуть простые гибриды диаллельному скрещиванию и испытать их на комбинационную способность. В полном объёме выполнить это нельзя, поэтому общее число возможных комбинаций для двойных гибридов определяют по формуле (8.7)
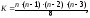 где
n
– число линий.
где
n
– число линий.
Для определения лучших двойных комбинаций можно использовать математический путь на основе данных диаллельного анализа (испытание на СКС). Эмпирически М. Дженкинс нашёл, что урожайность двойного гибрида имеет высокую корреляцию со средней урожайностью четырёх простых гибридов, не используемых в завершающем скрещивании.В частности из четырёх линий двойного межлинейного гибрида (А х В) х (С х Д) можно создать всего шесть простых гибридов: А х В, А х С, А х Д, В х С, В х Д, С х Д. Первый и последний из них родительские, все остальные – неродительские гибриды. Урожайность двойного гибрида можно приблизительно рассчитать по формуле (8.8)
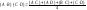 (8.8)
Математическое
определение урожайности гипотетических
двойных гибридов может служить только
для исключения из дальнейшей
селекционной работы тех комбинаций,
которые с высокой вероятностью не
относятся к высокоурожайным. Лучшие
комбинации испытывают на комбинационную
способность, чтобы найти действительно
лучшую двойную комбинацию.Современное
производство гибридных семян
основано на особенностях биологии
цветения. Если необходимо получать
гибридные семена для производства, то
ручная кастрация не применима. Мужская
стерильность
– явление, связанное с продуцированием
растениями пыльцы, не способной к
оплодотворению. Она возникает вследствие
полного недоразвития мужских
репродуктивных органов (пыльников) или
же в случае, когда в нормально развитых
пыльниках из-за нарушений мейоза не
образуются нормальные микроспоры и
пыльца становится нефункциональной,
или стерильной.Существует
три типа генетически обусловленной
мужской стерильности:
1) генетическая, или ядерная, – мужская
стерильность, обусловленная действием
одного и более генов в хромосомах; 2)
цитоплазматическая мужская стерильность
(ЦМС), обусловленная экстрахромосомными
факторами, или плазмогенами; 3)
цитоплазматически-ядерная мужская
стерильность, обусловленная действием
обоих факторов.Из всех типов стерильности
наиболее пригодной для селекции растений
оказалась цитоплазматическая мужская
стерильность.Цитоплазматическая
мужская стерильность основана на
взаимодействии между генами ядра
(хромосомами) и генами, находящимися в
цитоплазме. Причина
ЦМС заключается
в цитоплазме. Цитоплазму, содержащую
гены мужской стерильности, обозначают
символом S,
а цитоплазму, не обусловливающую мужскую
стерильность, – символом N.
Действие «стерильной» цитоплазмы
зависит от присутствия в ядре клетки
аллелей генов-восстановителей
фертильности, обозначаемых символами
Rf
– rf.
Стерильная цитоплазма S
вызывает стерильность мужских
генеративных органов цветка, если в
ядре клетки присутствует рецессивный
аллель rfгена-восстановителя
фертильности. Доминантный аллель
ингибирует (подавляет) действие
цитоплазматических генов мужской
стерильности. Таким образом, в зависимости
от генов, контролирующих мужскую
фертильность, возможны следующие типы
идиотипической конституции самоопылённых
линий:
(8.8)
Математическое
определение урожайности гипотетических
двойных гибридов может служить только
для исключения из дальнейшей
селекционной работы тех комбинаций,
которые с высокой вероятностью не
относятся к высокоурожайным. Лучшие
комбинации испытывают на комбинационную
способность, чтобы найти действительно
лучшую двойную комбинацию.Современное
производство гибридных семян
основано на особенностях биологии
цветения. Если необходимо получать
гибридные семена для производства, то
ручная кастрация не применима. Мужская
стерильность
– явление, связанное с продуцированием
растениями пыльцы, не способной к
оплодотворению. Она возникает вследствие
полного недоразвития мужских
репродуктивных органов (пыльников) или
же в случае, когда в нормально развитых
пыльниках из-за нарушений мейоза не
образуются нормальные микроспоры и
пыльца становится нефункциональной,
или стерильной.Существует
три типа генетически обусловленной
мужской стерильности:
1) генетическая, или ядерная, – мужская
стерильность, обусловленная действием
одного и более генов в хромосомах; 2)
цитоплазматическая мужская стерильность
(ЦМС), обусловленная экстрахромосомными
факторами, или плазмогенами; 3)
цитоплазматически-ядерная мужская
стерильность, обусловленная действием
обоих факторов.Из всех типов стерильности
наиболее пригодной для селекции растений
оказалась цитоплазматическая мужская
стерильность.Цитоплазматическая
мужская стерильность основана на
взаимодействии между генами ядра
(хромосомами) и генами, находящимися в
цитоплазме. Причина
ЦМС заключается
в цитоплазме. Цитоплазму, содержащую
гены мужской стерильности, обозначают
символом S,
а цитоплазму, не обусловливающую мужскую
стерильность, – символом N.
Действие «стерильной» цитоплазмы
зависит от присутствия в ядре клетки
аллелей генов-восстановителей
фертильности, обозначаемых символами
Rf
– rf.
Стерильная цитоплазма S
вызывает стерильность мужских
генеративных органов цветка, если в
ядре клетки присутствует рецессивный
аллель rfгена-восстановителя
фертильности. Доминантный аллель
ингибирует (подавляет) действие
цитоплазматических генов мужской
стерильности. Таким образом, в зависимости
от генов, контролирующих мужскую
фертильность, возможны следующие типы
идиотипической конституции самоопылённых
линий:
1. Мужская фертильность, обусловленная фертильной цитоплазмой – NRfRf, Nrfrf.2. Мужская фертильность, вызванная подавлением доминантного аллеля гена Rf стерильной цитоплазмы – SRfRf.3. Мужская стерильность – Srfrf.Ядерно-плазменные взаимоотношения, влияющие на фертильность мужских органов цветка, были найдены у кукурузы, сорго, проса, хлопчатника, табака и др.У кукурузы известны четыре формы ЦМС:
1. Техасский тип (Т) – экологически более стабильный, но связан с чувствительностью кукурузы к гельминтоспориозу.2. Молдавский тип (М), или S, или USAD-тип.3. С-тип. Источником данного типа ЦМС является образец из Бразилии, полученный в 1975 г. из США.4. Боливийский тип (В) пригоден только для производства простых гибридов, так как экологическая стабильность мужской стерильности у него слабая.Большинство гибридов получены на основе ЦМС техасского и молдавского типов. Из них большинство трёхлинейных, двойных и простых межлинейных.
44. Агробактериальная и биобаллистическая трансформация растений, примеры создания трансгенных сортов растений. Главные культуры, поставляющие более 60% пищи для человека и сельскохозяйственных животных - это злаки. За исключением риса трансформация злаков при помощи агробактерии в большинстве случаев не эффективна.Эффективная генетическая трансформация злаков возможна только при применении прямого способа ввода генов в клетки растений, опубликованного в 1988 году и названного авторами Джоном Стэнфордом и Теодором Клейном биобаллистическим (biolistic - биологическая баллистика). Суть его заключается в следующем. На микроскопические (размером с десятые доли микрон) вольфрамовые или золотые частицы наносится молекула ДНК, содержащая нужные гены. В специальной вакуумной камере металлические частицы, покрытые ДНК, разгоняются до скоростей, достаточных для проникновения в клетки-мишени, компетентные к регенерации растений in vitro. Здесь уместно пояснить несколько терминов, широко используемых в генной инженерии. Генетическая трансформация - процесс введения чужеродного гена в растение, или получения растения с работающим и передающимся по наследству чужеродным геном; экспрессия - работа гена, завершающаяся синтезом белка, который данный ген кодирует; вектор - молекула ДНК, содержащая ген, который необходимо встроить в геном растения, и последовательности ДНК, ответственные за процесс встраивания и экспрессию данного гена; in vitro - означает "в стекле", то есть в пробирке; каллус - неорганизованно растущая масса клеток в пробирке; регенерация растений in vitro - получение целого растения из каллуса или отдельных клеток в пробирке.Клетки растения помещаются в вакуумной камере под источником частиц, и в ходе "обстрела" гены доставляются в клетки и встраиваются в геном растения. Затем следует этап селекции трансформированных клеток и регенерации трансгенных растений in vitro.
Поэтому для отбора пригодных для селекционного процесса генетически модифицированных форм необходимо иметь, как показывает практика, несколько десятков или даже сотен растений, полученных от независимых трансгенных событий. Столь большую выборку можно получить только при высокой эффективности трансформации. Для трудно регенерируемых in vitro злаков и подсолнечника реальный выход получаемых трансгенных растений составляет менее 1-2%.Создание трансгенных растений в настоящее время развиваются по следующим направлениям:1. Получение сортов с/х культур с более высокой урожайностью 2. Получение с/х культур, дающих несколько урожаев в год (например, в России существуют ремантантные сорта клубники, дающие два урожая за лето) 3. Создание сортов с/х культур, токсичных для некоторых видов вредителей (например, в России ведутся разработки, направленные на получение сортов картофеля, листья которого являются остро токсичными для колорадского жука и его личинок) 4. Создание сортов с/х культур, устойчивых к неблагоприятным
климатическим условиям (например, были получены устойчивые к засухе
трансгенные растения, имеющие в своем геноме ген скорпиона) 5. Создание сортов растений, способных синтезировать некоторые белки животного происхождения (например, в Китае получен сорт табака синтезирующий лактоферрин человека)