

13. Автономна (вегетативна) нервова система
Усі функції організму можна умовно поділити на дві групи: соматичні (анімальні, тваринні) і вегетативні (вісцеральні, рослинні) (від лат. vegetativus-рослинний). Такий поділ запропонував у 1801 р. М.Біша.
До соматичних відносять сприймання зовнішніх подразнень і рухові реакції поперечно-смугастих м'язів, а до вегетативних - процеси, що забезпечують обмін речовин, травлення, дихання, кровообіг, виділення, розмноження тощо.Відповідно до розподілу функцій нервову систему також поділяють на соматичну і вегетативну.У 1903 році Д.Ленглі ввів поняття «автономна нервова система», підкреслюючи, що її функціонування майже не залежить від нашої свідомості, і запропонував поділ на симпатичну і парасимпатичну частини (в останній час виділяють ще метасимпатичну частину).У 1916 році Гаскел запропонував термін «мимовільна».Згідно міжнародній анатомічній номенклатурі, термін автономна замінив усі раніше існуючі -рослинна, вісцеральна, вегетативна, хоча насправді цей відділ нервової системи не є повністю автономним.В поняття автономна нервова система (АНС) включається комплекс центральних і периферичних нервових клітинних структур, котрі регулюють функціональний рівень внутрішнього середовища, необхідний для адекватних реакцій всіх систем. Досягається це шляхом іннервації гладеньких м'язів усіх органів, серця і залоз.
13.1. Структурно-функціональні особливості автономної (вегетативної) нервової системиАНС має слідуючі відмінності порівняно з соматичною.
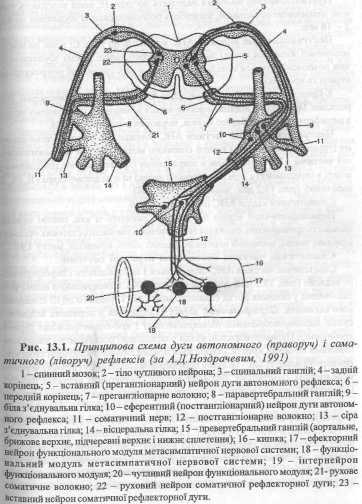
1. Еферентний відділ рефлекторної дуги АНС має два нейрони - ефек-торний і вставний (рис. 13.1).
Ефекторний нейрон розташований поза межами ЦНС і входить до складу одного з вузлів АНС. Вставний нейрон міститься в середині мозку, внаслідок чого зветься центральним. Саме ця властивість вважається принциповою ознакою вегетативної нервової системи на відміну від соматичної.
2. Еферентні волокна, які виходять з центрів АНС, на шляху до органа перериваються в тих чи інших вузлах і відповідно називаються преганглі-онарпими і постгангліонарними.
132
Прегангліонарні волокна належать до типу В; еони володіють тонкою мієліновою оболонкою, швидкість розповсюдження збудження по них 3-18 м/с.
Постгангліонарні волокна АНС належать до типу С; вони не мають мієлінової оболонки, тонкі (діаметр їх не перебільшує 7 мкм), збудження по них розповсюджується повільно (1-3 м/с), у них триваліші рефрактерний період та хронаксія.
3. Центральні й периферичні відділи АНС розміщені нерівномірно, осередками. Центральні відділи локалізуються в спинному й головному мозкч, а периферичні мають багато нервів, нервових сплетень і вузлів.
Центре сямпатичної частини АНС розташовані в торако-люмбально-му (грудно-поперековому) відділі спинного мозку. Центри парасимпатичної частини АНС розташовані в сакральному (крижовому) відділі спинного мозку, в середньому (в ядрі Ш черепного нерва) та довгастому (ядраУП, ІХ. X пар) мозку (рис. 13.2 див. додаток на стор. 208).
Симпатичний відділ АНС іннервує все тіло.
Парасимпатичний відділ не іннервує скелетні м'язи, потові залози, більшість кровоносних судин (за виключенням зовнішніх статевих органів і слинних залоз), мозковий шар надниркових залоз, сечоводи, матку, ЦНС.
Симпатична частина автономної нервової системи
Симпатична частина АНС складається з центрального і периферичного відділів (рис. 13.3 див. додаток на стор. 209). Центральний відділ розташований у бокових рогах спинного мозку від І грудного до Ш поперекового сегмента. Периферичний відділ складається з нервових волокон і вузлів паравертебральних (біжхребтових) і превертебральних (передхребтових). Паравертебральні вузли розташовані посегментно двома ланцюжками по боках хребта, утворюючи правий і лівий симпатичні стовбури. Превертебральні вузли - це вузли периферійних сплетень у грудній І черевній порожнинах (аортальне, брижове верхнє, підчеревні верхнс і нижнє). Симпатичні нервові волокна виходять зі спинного мозку в складі передніх корінців спинномозкових нервів, а потім через прегангліонарні (передвузлові) волокна - білу сполучну гілку - прямують до відповідного вузла симпатичного стовбура. Там частина волокон переходить на постгангліонарний (післявузловий) нейрон, і до органів доходять його волокна. Друга частина проходить через вузол симпатичного стовбура не перериваючись, і підходить до превертебральних вузлів, перемикається в них, а звідти до органів тягнуться постгангліонарні волокна.
Симпатичний стовбур - правий і лівий - є ланцюжком нервових вузпів уздовж хребта, з'єднаних міжвузловими гілками. Шийний відділ має три вузли- верхній, середній і нижній. Нижній шийний яерідко зливається з верхнім грудним, утворюючи шийно-грудний (зірчастий) вузол. Від верхнього шийного вузла відходить внутрішній сонний нерв, який іннервує слізну залозу, слинні залози та м'яз, що розширює зіницю, м'язи піотки і гортані, Від усіх трьох шийних вузлів симпатичного стовбура відходять серцеві нерви, які разом з блукаючим нервом проникають у стінку серця. Від грудних вузлів відходять гілки до аорти, серця, легенів, бронхів, стравоходу, де утворюють органні сплетення.
Частина прегангліонарних волокон, пройшовши транзитом вузли симпатичного стовбура, утворюють великий і малий нутряні нерви. Обидва нерви проходять через діафрагму в черевну порожнину, де частина з них закінчується у вузлах черевного (сонячного) сплетення. Друга частина опускається в малий таз і доходить до нейронів підчеревного сплетення. Поперековий відділ симпатичного стовбура складається з 3-5 вузлів, гілки від яких беруть участь в утворенні сплетень черевної порожнини й таза.
Черевне сплетення (ріехия соеііасш) - найбільше в АНС, міститься між наднирковими залозами. Його утворюють парні черевні вузли і непарний верхній брижовий вузол. їхні післявузлові волокна утворюють черевне аортальне сплетення, яке іннервує надниркові і статеві залози, нирки, шлунок, підшлункову залозу, селезінку, тонку і товсту кишки до низхідної ободової {верхнє брижове сплетення). Нижнє брижове сплетення розташоване на аорті; проходячи вздовж гілок нижньої брижової артерії, дає розгалуження в низхідну ободову, сигмоподібну й верхню частину прямої кишки.
Підчеревні сплетення (верхнє і нижнє) розташовані на черевній аорті. Післявузлові волокна сплетення розходяться по гілках внутрішньої клубової артерії й іннервують нижні відділи прямої кишки, сечовий міхур, сім'явиносну протоку, передміхурову залозу, матку і піхву.
Існує думка, що, окрім еферентних волокон, симпатична нервова система має власні чутливі шляхи. Залежно від локалізації клітинних тіл, ходу й довжини гілок вони можуть бути поділені на дві групи. Перша група периферійних аферентних нейронів включає клітини, тіла яких локалізуються в превертебральних симпатичних вузлах. Одна з довгих гілочок прямує на периферію, інша - у бік спинного мозку, куди входить у складі дорзальних корінців. Друга група характеризується тим, що довга гілочка цих чутливих ^гїїин пов'язана з робочим органом, короткі ж розподіляються в самому
вузлі, синаптично контактують із вставними нейронами, а через них - з ефек-торними нейронами й створюють тут місцеву рефлекторну дугу. Парасимпатична частина автономної нервової система
Парасимпатична частина АНС також має центральний і периферичний відділи. Центральний відділ складається з парасимпатичних ядер, закладених у середньому та довгастому мозку й крижових сегментах (П -IV) спинного мозку. Периферичний відділ - це вузли й волокна, що входять до складу окорухового (ПІ пара), лицевого (VII пара), язикоглоткового (IX пара), блукаючого (X пара) і тазових нервів.
У середньому мозку на дні водопроводу лежить парасимпатичне додаткове окорухове ядро (ядро Якубовича-Едінгера-Вестфаля), відростки клітин якого прямують у складі окорухового нерва, переключаються у війковому вузлі (міститься в очній ямці) і закінчуються в м'язі, який звужує зіниці, та у війковому м'язі.
У ромбоподібній ямці поряд з ядром лицевого нерва містяться слиновидільне черепне (верхнє) ядро. Відростки його клітин входять до складу проміжного нерва, потім лицевого. Разом із гілками лицевого, ата-кож трійчастого нервів парасимпатичні волокна досягають слізної залози, залоз слизової оболонки носової і ротової порожнин (перемикаються в крилопіднебінному вузлі) та піднижньощелепної і під'язикової залоз (перемикаються у розташованому поряд піднижньощелепному вузлі).
Слиновидільне хвостове (нижнє) ядро дає початок парасимпатичним (секреторним) волокнам привушної залози, які виходять з мозку в складі IX пари (язикоглоткового нерва) і перемикаються у вушному вузлі.
Основна маса парасимпатичних волокон, які виходять з довгастого мозку, входять до складу блукаючого нерва. Вони починаються від його парасимпатичного спинного (дорзального) ядра на дні ромбоподібної ямки. Передвузлові волокна тягнуться до органів шиї, грудної та черевної порожнин, закінчуючись в інтрамуральних (всередині органів) вузлах щитоподібної та вилочкової залоз, у бронхах, легенях, серці, стравоході, шлунку, кишках (до селезінкового вигину), у підшлунковій залозі, печінці, нирках. Від інтрамуральних вузлів відходять післявузлові волокна, які іннервують ці органи.
Від крижових сегментів спинного мозку парасимпатичні передвузлові волокна прямують у складі вентральних корінців крижових нервів, а відділившись від них, утворюють нутряні тазові нерви. їхні гілки входять до підчеревного сплетення і закінчуються на клітинах інтрамуральних вузлів.
є Післявузлові волокна іннервують гладкі м'язи і залози нижніх відділів трав-I ного апарату, сечовидільні, зовнішні й внутрішні статеві органи. Ш Головним колектором чутливих шляхів парасимпатичної нервової Ц системи є блукаючий нерв. Аферентні волокна його шийного відділу скла->- дають 80 - 90%. Приблизно 20% з них є міелінізованими, регата - тонкі 1. безміелінові. Ці волокна передають інформацію від травного тракту, органів : грудної та черевної порожнин. Створені цими волокнами рецептори реагу-■. ють на механічні, термічні, больові впливи, сприймають зміни рН і ,д електролітного складу.
І. Надзвичайно важлива фізіологічна роль чутливої гілки блукаючого не-к.рва - депресорного нерва. Він є потужним провідником, що сигналізує про Шрівень артеріального тиску в аорті. Клітинні тіла аферентних шляхів блука-
■цочого нерва розташовано здебільшого в яремному вузлі, а їх волокна всту-
Иьають у довгастий мозок на рівні олив.
Щ! У складі синусного нерва, що є гілочкою IX пари, міститься близько
^300 товстих аферентних волокон, які зв'язані з великою кількістю рецепторів ^різної модальності. У цьому сприймаючому комплексі особлива роль належить каротидному клубочку, який розташовано між внутрішньою і ^зовнішньою сонними артеріями в місці поділу спільної сонної артерії (сонній пазусі, 5ІШ15 сагоіісиз).
IV 13.2. Метасимпатична нервова система
■,-. Багато внутрішніх органів після перетинання симпатичних і парасимпа-тичних шляхів, або навіть після вилучення з організму, продовжують
^здійснювати властиві їм функції без особливих змін. Зберігає координовану перистальтичну й всмоктувальну функції кишка, скорочується серце. Сегменти або смужки матки, сечоводу, жовчного міхура продовжують скорочуватися з частотою й амплітудою, що властива кожному органу. Ця функціональна автономія пояснюється наявністю в стінках органів, яким притаманна автоматія, гангліозної системи, котра здійснює регуляторний вплив на них. Для здійснення автономної рефлекторної та інтегративної діяльності є всі ланцюги - чутливі, вставні, рухові, а також медіатори. Цю систему виділено в метасимпатичну части-
,нуАНС(рис. 13.4).
Отже, більшість порожнистих вісцеральних органів, поряд з екстраор-ганпою симпатичною і парасимпатичною нервовою системою, мають впас-
ний базовий механізм нервової регуляції. Керування роботою в цьому випадку забезпечується рефлекторними дугами, що замикаються в межах стінки самих органів.
Метасимлатична нервова система володіє багатьма ознаками, які відрізняють її від інших частин АНС.
• Іннервує тільки внутрішні органи, які володіють власною моторною активністю, тобто автоматіею; у сфері її іннервації знаходяться гладкі м'я-зн, всмоктувальний і секретуючий епітелій, локальний кровообіг, місцеві ендокринні елементи.
• Одержує синаптичні входи від симпатичної та парасимпатичної нервової системи й не має прямих синаптичних контактів з еферентною частиною соматичної рефлекторної дуги.
• Поряд із загальним вісцеральним аферентним шляхом має власний сенсорний ланцюг.
• Не знаходиться в антагоністичних відносинах з іншими частинами нервової системи, але, являючи собою істинно базову іннервацію, володіє
Значно більшою, ніж симпатична й парасимпатична нервова системи, незалежністю від ЦНС.
• Органи з пошкодженими або з вимкненими за допомогою гангліоблокаторів метасимпатичними шляхами втрачають властиву їм здатність до координованої ритмічної моторної функції.
• Метасимпатична нервова система володіє власним медіаторним ланцюгом.
Отже, метасимпатична система звільняє ЦНС від надмірної інформації та разом з тим забезпечує надійність регуляції функції. Метасимпатична система- це своєрідний мікропроцесор, розташований поблизу від ефек-дарів (гладеньких м'язів, екскретуючого, всмоктуючого епітелію, ендокринних елементів). Апарат метасимпатичної системи ініціює та програмує роботу ефекторІв, здійснюючи біжучу регуляцію заради підтримання постійності внутрішнього середовища. Поряд із цим, АНС (разом з іншими відділами ЦНС) діє як головний комп'ютер, виконує призначення стратегічного керування в якості модулятора, особливо на початку і в кінці дії. У ЦНС записано певні програми для забезпечення сенсомоторної регуляції та координації ефекторних структур цілісного організму.
13.3. Рефлекторні дуги та центри автономної нервової системи
Чутлива ланка дуги автономного рефлексу
Рецептори вісцеральних органів (інтерорецептори) за будовою поділяють на вільній невільні. Тнтерорецептори здатні сприймати механічні й хімічні стимули.
Механорецептори часто звуться барорецепторами, їх поділяють на два типи: такі, що повільно й такі, що швидко адаптуються.
Хеморецептори поділяють на кислото- і лужночутливі.
Інтерорецептори розташовуються в стінках практично всіх вісцеральних органів: судинах, серці, легенях, шлунку, кишечнику, нирках, сечовому міхурі, печінці та ін.
Реакція рецептора на стимул складає основу для послідуючого аналізу сенсорних впливів, який здійснює нервова система. Саме від рецепторів залежить межа чутливості й діапазон впливів, що сприймаються.
Чутливі закінчення, завдяки анатомічним і функціональним особливостям, вибірково реагують на один певний вид енергії. З другого боку, сти-

мул, яким би він не був за своєю природою, -механічна деформація, хімічні зміни, температурне зрушення - завжди викликає один і той же електричний сигнал (потенціал дії), який відіграє роль символа. Інформація про інтенсивність стимула кодується частотою імпульсів. Важливо й те, що рецептори не тільки сприймають, але й підсилюють сигнал, який сприймають.
Нервові волокна, по яких вісцеральна сенсорна інформація передається до центрів інтеграції, поділяються на групи А, В, С. На відміну від метамерності, з якою в спинний мозок посипають інформацію соматичні аференти, провідники вісцеральної чутливості мають складний полісегментарний розподіл, що створює умови для мультиплікації їх дії.
При надходженні до спинного мозку частина аферентних волокон первинно зв'язується з сегментарними інтеряейронами, які, в свою чергу, си-наптично контактують із проміжними або еферентними клітинами того ж чи ближніх сегментів. Так створюється дуга автономного рефлексу спи-пального рівня. Друга частина аферентних волокон розподіляється слідуючим чином: висхідні гілки частково закінчуються в сірій речовині розташованих вище сегментів, частково досяі-ають дорзальної частини довгастого мозку, де й закінчуються в ядрах задніх стовпів. Тут розташовано другі нейрони аферентного шляху, аксони яких формують бульботаламічнтш шлях. Низхідні гілочки опускаються на 6 - 7 см і утворюють синаптичні контакти з асоціативними клітинами розташованих нижче сегментів.
Аферентні волокна можуть бути звичайними задньокорІнцевими й належати як соматичній, так і вегетативній рефлекторній дузі сішнального рівня. Слідуюча ланка цієї ж дуги є представленою інтернейроном спинного мозку, який синаптично зв'язується з нейроном бокових рогів. Отже на спи-нальному рівні має бути не менш як два синаптичнях перемикання.
Існують міркування й на користь своєрідних "симпатичних" аферентних волокон. По-перше, то волокна клітин розташованих в превертсбраль-ному симпатичному вузлі, периферичний паросток яких закінчується рецепторами, а центральний завершується на еферентному нейроні в самому ганглії (утворюючи "місцеву" рефлекторну дугу), або через білу сполучи)' гілку надходить до спинного мозку.
Нарешті, варіант, коли клітина аферентного нейрона розташована на периферії, у складі функціонального модуля метасимпатичної системи, а аферентне волокно підіймається доцентрово аж до спинного мозку чи вегетативного ганглія.
Асоціативна (вставна) ланка
У дузі автономного рефлексу перемикання аферентних сигналів на асоціативні й далі на еферентні клітини може відбуватися в периферійних утвореннях - в превертебральних та інтрамуральних гангліях, так званих . нижчих центрах, а також на сгашальному Й бульбарному рівнях. ■: Тонка координація діяльності всіх трьох частин АНС здійснюється над-" Сегментарними центрами різних рівнів, аж до кори великих півкуль головного мозку.
Сііивальні механізми. У спинному мозку існує спеціальний
інтернейронний апарат, який здійснює первинне опрацювання аферентних
сигналів. Цей апарат з'єднаний з клітинами бокових рогів спинного мозку.
Така організація, мабуть, узгоджує аферентні сигнали, які надходять одно-
ІІчасно від різних рецепторних ділянок. Інтеграція сигналів не обмежується сег-
^Ьентарним спинномозковим рівнем і в певних умовах може здійснюватися
?|Йвнямивищого порядку.
Таким чином, у дузі автономного рефлексу між чутливим ї , постгангліонарними нейронами розташовано не менше трьох синапсів, два іі яких знаходяться в сірій речовині спинного мозку, а один - в периферійному автономному ганглії.
Існування різних власних вихідних елементів в асоціативній ланці спи-
нального рівня, можлива присутність спеціалізованих збуджуючих
інтернейронів, гальмівних механізмів і низхідних симпатоактивуючих шляхів
роблять цілком виправданим введення В.П.Лебєдєвим для позначення цієї
^Йнстеми спеціального поняття симпатичний механізм спинного мозку.
%
', Суцраспивальяі механізми. Подальше опрацювання аферентних сигналів вісцеральної чутливості, що надходить по провідникових шляхах спи?і-ного мозку, відбувається в ретикулярних утвореннях заднього мозку, корі мозочка, вестибулярних ядрах та інших структурах. Крім того, тут взаємодіють сигнали, які надходять і по аферентних волокнах черепних нервів. Взаємодії аферентних вісцеральних і соматичних сигналів активуються ретикулярними нейронами, які надсилають низхідні сигнали до спинальних соматичних і прегангліонаряих нейронів дуги автономного рефлексу.
По внутрішньоцентральних висхідних шляхах вісцеральні потоки
Ї>ів досягають середнього мозку, ретикулярна формація якого є сут-нстанцією на шляху до асоціативних ділянок кори великих півкуль. * - . ■ "' ЙГ
Особливістю представництва є те, що в одній і тій же точці реєструються відповіді не тільки від вісцеральних, але й соматичних чутливих закінчень.
Для кожної з аферентних систем, за виключенням нюхової, у проміжному мозку є спеціальне перемикаюче первинне «реле» з високим ступенем диференціювання й щільністю представництва. На цьому рівні можуть здійснюватися не тільки різні перемикання на вісцеральні рефлекторні системи, але й взаємодія між різними рефлекторними сигналами. До структур проміжного мозку, що здійснюють цю функцію, відносяться специфічні їі неспецифічні ядра таламусу, а також гіпоталамічні структури.
Зона представництва головного колектора чутливих шляхів черевної порожнини (черевного нерва) розташовується в латеральній частині заднь-овентрального ядра таламусу. Блукаючий нерв представлений в медіальній зоні цього ж ядра. Ділянки представництва кожного з цих нервів перекриваються проекціями соматичних аферентів: черевного - від тулуба, блукаючого - від плечового пояса. Це означає, що в проекційних ядрах таламусу відбувається взаємодія Інформації, що надходить по вісцеральних і соматичних аферентних шляхах.
Кортикальне представництво. В.Я.Данилсвський в 1874 р. установив, що подразнення деяких ділянок кори великих півкуль у собак викликає зміни дихання, серцевої діяльності, тонусу судин.
Представниками школи К.М.Бикова зформульовано ствердження про те, що кора великих півкуль приймає участь у регуляції функцій внутрішніх органів, тобто про коркове представництво інтероцептивного аналізатора.
В організації проекцій аферентних систем у корі великих півкуль виділено два типи представництва: первинне і вторинне. Первинне пов'язане з локальним розподілом в одній ділянці кори переважно одного типу чутливих зв'язків. Особливістю вторинного представництва є дифузний розподіл проекцій аферентних систем. Вони реєструються за межами первинних проекційних ділянок (В.Н.Чершгівський).
Чутливі імпульси, що прямують по черевних нервах, проекуються контра- та іпсилатерально в І і П соматосенсорні ділянки. Тут також представлено соматичну чутливість. На цій основі вважають, що об'єднання сигналів вісцеральної та соматичної природи відбувається не тільки в таламічних ядрах, але й на рівні кори мозку.
імпульси, що прямують від шийної, грудної й піддіафрагмальної частин блукаючого нерва, реєструються в орбітальній, поясній закрутках.
Не дивлячись на складність багатоступеневої організації, основа механізму міжнейронної взаємодії на всіх рівнях та ж сама - синаптична конвергенція на центральних клітинах сигналів різної природи (вісцеральної й соматичної), їх оцінка за принципом сумації й на основі результатів оцінки - генерація нового сигналу.
Значення кори великих півкуль головного мозку в регуляції функцій органів, що іннервуються АНС, і роль останньої як провідника імпульсів від кори великих півкуль до периферичних органів яскраво виявляються в дослідах з умовними рефлексами на зміни діяльності внутрішніх органів.
Еферентна ланка
Еферентна ланка дуги автономного рефлексу являє собою нервову клітину, котра мігрувала зі спинного мозку в одинз периферичних гангліїв. Ганглії автономної нервової системи є нижчими інтегративними центрами.
Прегангяюнарні нейрони одержують входи від периферичних вісцеральних І соматичних чутливих клітин, а також по низхідних шляхах від вищих центрів.
Підходячи до ефекторних клітин, прегангліонарне волокно втрачає мієлін І, розгалужуючись, створює на тілі й відростках постгангліонарних клітин синаптичиі контакти. Прегангліонарних волокон значно менше ніж постгангліонарних нейронів. Так, до верхнього шийного симпатичного вуз-лау людини підходить 10 тис. прегангліонарних волокон, які закінчуються На мільйоні постгангліонарннх клітин вузла. Таким чином, одне Ярегангліонарне волокно контактує не менш ніжзі ста постгангліонарними нейронами. Крім того, на тому ж самому постгангліонарному нейроні може шввергувати декілька прегангліонарних волокон. Така широка конвергенція йдивергенція забезпечує високу надійність проведення збудження в гангліях. Завдяки дивергенції здійснюється у вегетативних гангліях феномен мультиплікації. Полягає він у тому, що одна й та ж інформація, яка надходить з Одного центру (від одного нейрона) копіюється й сповіщається через си-Напси багатьом нейронам вегетативного ганглію, а через постгангліонарні волокна - багатьом виконавчим органам. Цим забезпечується принцип економії, тобто розвантаження нервових центрів від зайвої маси нейронів. Вегетативний ганглій виконує функцію нервового центру, тобто с «центром», винесеним на периферію, за межі ЦНС. По-друге, у такий спосіб внутрішні Органи (та їх частини) практично ніколи не активуються ізольовано; у системній діяльності органів завжди підтримується узгоджена одночасність.
У парасимпатичних вузлах така конвергенція практично відсутня. Кожне прегангліонарне волокно контактує тільки з однією-двома постганппіонарними клітинами. Цим пояснюється той факт, що парасимпатична стимуляція завершується, як правило, менш поширеною, ніж симпатична, відповіддю.
Чітко виражена конвергенція на нейронах вегетативних гангліїв закінчень прегангліонарних волокон забезпечує інтенсивне перекриття зон впливів від нейронів різних, вегетативних центрів і гангліїв, як передумова для інтегративних процесів обробки інформації за принципом просторової та часової сумації, трансформації ритму, оклюзії та полегшення.
Велика тривалість електричних процесів - ЗПСП, ЦД (у значній мірі завдяки слідовій гіперполяризації) і відповідно велика тривалість фаз реф-рактерності є причиною низької лабільності у вегетативних синапсах і отже низької частоти імпульсів, які надходять до робочих органів. Мабуть з цим пов'язана й така особливість проведення імпульсів через вегетативний си-напс як ітсративність, тобто необхідність для проведення збудження через синапс серії імпульсів. Існує думка, що сповільнення процесів переробки інформації в гангліях пов'язане з наявностю в їх складі певних інтерней-ронів - вставних катехоламінергічних. До речі, у вегетативних синапсах триваліша дія медіаторів за рахунок меншої активності ферментів, які їх розщеплюють.
Взаємодіючи з метасимпатичною нервовою системою, прегантліонарні парасимпатичні й постгангліонарні симпатичні волокна модулюють активність виконавчого вісцерального органу. При цьому припускається, що парасимпатичні волокна здатні збуджувати не тільки еферентні нейрони метасимпатичної нервової системи, що активують, наприклад, моторну діяльність кишки, але й гальмівні клітини. Симпатичні ж впливи, поряд зі слабкою прямою дією на гладкі м'язи органа, викликають виникнення гальмівних процесів у холінергічних нейронах місцевих метасимпатичних сіток, а також гальмують вихід медіатора з прегангліонарних парасимпатичних волокон. Таким чином, модулюючи приплив імпульсів по парасимпатичних шляхах, симпатична нервова система тим самим надає регулюючий вплив на моторну активність таких органів, як кишка і сечовий міхур.
Еферентний нейрон може мати декілька входів. Він володіє рядом морфологічних і функціональних властивостей, які дозволяють інтегрувати вхідні сигнали і створювати модифікований вихідний сигнал. Головний вхід - збуджуючий, холінергічнии - представлено прегангліонарними волокнами. Слідуючий вхід виключно периферичного походження. Тіла цих чутли-
144
ь і і - і шйронів можуть розташовуватись або в самому ганглії, у безпосередній близькості від еферентного нейрона, або в гангліях стінок внутрішніх органів, тобто мають метасимпатичиу природу. Завдяки цим місцевим рефлекторним дугам в еферентному нейроні підтримується необхідний рівень спонтанної активності й зберігається рефлекторна діяльність при децентралізації вузла.
Частина еферентних нейронів спонтанно активна. У деяких з них ритм і патерн розряду співпадає з показниками активності прегангліонарних волокон або відбиває приплив по вісцеральних волокнах місцевих рефлекторних дуг. Здебільшого відзначається кореляція «входів» і «виходів». Фонова активність еферентних нейронів низькочастотна, розряди клітин можуть з'являтися в момент серцевого поштовху, фази вдиху або синхронно з пе-ристальтич цими рухами кишки.
13.4. Рефлекси в залученням ланок автономної рефлекторної дуги
К Процеси в автономній і соматичній системах тісно пов'язані, хоча при Вгодразненні автономних чутливих волокон обидві системи залучаються до ріідповіді в різному ступені. Рефлекси в цьому випадку поділяються на \ісцеро - вісцеральні, вісцеросоматичні, вісцеросенсорні.
Вісцеро - вісцеральний рефлекс обумовлений шляхами, у котрих збудження виникає й завершується у внутрішніх органах. Прикладами вісцеро-вісцеральних рефлексів можуть бути: класичний дослід Гольца, який ілюструє сповільнення серцевих скорочень при механічному подразненні брижі; зміни артеріального тиску, частоти серцевих скорочень при подразненні каротидної й аортальної рефлексогенних зон.
Основою для здійснення цих процесів можуть бути й місцеві рефлекторні Дуги.
Вісцеросоматичпий рефлекс забезпечується шляхами, збудження яких, на доповнення до вісцеральних, викликає також соматичні відповіді. Відомо, що подразнення хемо- і механорецепторів каротидної зони супроводжується гальмуванням загальної рухової активності тощо.
Вісцеросснсорний рефлекс забезпечується шляхами, у котрих у відповідь на подразнення автономних чутливих волокон виникають реакції не тільки у внутрішніх органах, м'язах, але й змінюється соматична Чутливість.
Соматовісцеральннй рефлекс - активація вісцерорецепторів і сома-
І
145
тичних аферентних волокон викликає зміни у вісцеральних органах. Прикладом можуть бути реакції судин при термічних впливах на рецептори шкіри.
З наявністю спряжених рефлексів між вісцеральною й соматичною системами ми зустрічаємось у клініці. У разі патологічної інтероцептивної імпульсації від ураженого органу виявляються вісцеро-дермальні рефлекси, як наслідок аферентації від патологічного вогнища. Проявами можуть бути локальне потовиділення, почервоніння та підвищена електропровідність шкіри (відповідно із зонами Захар'їна-Геда).
Класичною ознакою вісцсро-моторного рефлексу є захисне напруження м'язів черевного пресу (іІеГепзе) у ділянці проекції ураженого запальним процесом органу з відповідною частиною перітонеуму. До такої ж категорії захисних рефлексів можна віднести й певну вимушену позу. Прикладом соматовісцерального рефлексу є знеболююча дія теплої грілки на шкіряну зону, де спроектовано уражений орган.
13.5. Медіатори автономної нервової системи
Особливості й механізми синаптичної передачі в автономній нервовій системі принципово такі ж, як і в соматичній. Однак в автономній нервовій системі пре- і постсинаптичні утворення значно різноманітніші. Це нейрони, клітини гладких м'язів, залоз та міокарду (рис. 13.5). Широко варіює також щільність автономної іннервації різних органів.
У сім'явиносній протоці, у війковому тілі відстань між розширенням аксону й мембраною м'язової клітини складає близько 20 нм. Тому тут м'язові клітини повністю підкоряються нервовому контролю й катехоламіни крові на них не впливають. У більшості ж кровоносних судин відстань між розширенням аксону й мембраною м'язової клітини складає більше 80 нм. Унаслідок такої організації на м'язи судин впливають катехоламіни крові,
В АНС нараховується більше десятка медіаторів - ацетилхолін, норадреналін, серотонін та інші біогенні аміни, АТФ і амінокислоти. У зв'язку з цим нейрони, що іх утворюють, називають холінергічними, адренергічними, серотонінергічними, пуринергічними, і т.п. Існує також багато типів гіпоталамічних нейронів, які продукують нейрогормони.
Ацетилхолін
Ацетилхолін (Ах) звільняється б закінченнях усіх прегангліонарних (і симпатичних, і парасимпатичних) нейронів і більшості постгангліонарних.
146

парасимпатичних нейронів. Крім того, частина постгангліонарних симпатичних нейронів, які прямують до потових залоз, і, можливо, симпатичні нейрони, що викликають розширення судин скелетних м'язів, також здійснюють передачу за допомогою Ах. У тканинах є фермент, що руйнує Ах - ацєтилхолінестераза.
Дія Ах може бути відтворена за допомогою фармакологічних препаратів. Так, нікотин викликає подібний ацетилхоліну ефект при дії на постсинаптичну мембрану постгантліонарного нейрона, токсин мухомора мускарин - на мембрану ефекторного органа. Така різниця в реакціях стала основою для виділення двох типів холінорецепторів: нікотинового (И-холінорецєптора) і мускаринового (М - холінорецептора).
Фармакологічні препарати, які впливають на ефекторний орган аналогічно постгангліонарному парасимпатичному нейрону, одержали назву парасимпатоміметиків.
М-холіноміметики (збуджують М-холінорецептори): пілокарпін, ацеклідип.
Н-холіноміметики (збуджують Н-холінорецептори): цитизин. Антихолінестеразні (блокують холінестеразу, при цьому ацетилхолін. який виділяється, не руйнується і Його ефект підсилюється й пролонгується; ефективніше діють на органи з М-хопінорецепторами); прозерин, оксазил - діють переважно на нервово-м'язову передачу; фосфакол, езерин - діють на ЦНС.
Існують речовини (холінолітики), які вибірково блокують синзптичну передачу в холінергічних синапсах.
Блокатори М-холінорецепторів: атропін і скополамін. Блокатори Н-холінорецепторів: а) гангліоблокатори - бензогексоніп, пентамгщ б) курареподібні (м'язові релаксанти, взаємодіють з Н-холіно-рецепторами скелетних м'язів) - тубокурарина хлорид, диплацин, дитиній.
Окрім медіаторної ролі, Ах володіє і загально-біологічною дією. Він пригнічує роботу серця, підсилює легеневу вентиляцію, скорочує м'язи бронхів, підсилює шлунково-кишкову перистальтику, активує секрецію травних залоз. Унаслідок короткочасності ефекту сам Ах не вживається, але використовуються його похідні метахолін і карбамілхолін.
Норадреналін і адреналін
Норадреналін відрізняється від адреналіну відсутністю метилового радикалу в бічному ланцюгу. Норадреналін є медіатором у постгангліо нарних симпатичних закінченнях судин серця, печінки, селезінки. Його вміст досягає 10% від загальної кількості катехоламінів у ділянках симпатичної
Іннервації. У мозковому шарі надниркових залоз, клітини яких гомологічні , постгангліонарному симпатичному нейрону, вміст норадренапіну досягає 20%, решта припадає на адреналін. У тканинах є фермент моноамінокси-.даза, який руйнує адреналін. Є ще й третій природний катехоламін у тканинах - дофамін.
Потрапляючи на постсинаптичну мембрану, норадреналін взаємодіє з її специфічними хімічними компонентами - адренорецепторами. На адре-иорецептори може діяти не тільки медіатор, але й аналогічні фармакологічні речовини, які називають адреноміметичними.
( Відповідно з чутливістю до різних катехоламінів адренорецептори поділяють на ά-адренорецептори та β-адренорецептори.
У більшості вісцеральних органів знаходяться обидва види адренорецепторів, а результати їх збудження бувають, як правило, проти-яежними. Наприклад, у гладких м'язах артерій скелетних м'язів є ά- і β- адренорєцептори. Збудження ά-адренорецепторів призводить до звуження артеріол, а збудження β-адренорецепторів - до їх розширення. У кишках також є обидва види адренорецепторів, однак вплив на ті та інші викликає гальмування активності гладких м'язів.
У серці й бронхах немає ά-адренорецепторів, і тут норадреналін і адреналін збуджують тільки β-адренорецептори (відповідно β1 та β2), що призводить до вазоділятації судин серця і до підсилення його скорочень та розширення бронхів.
Норадреналін, як і ацетилхолін, викликає ефект не тільки на постсинаптичній мембрані, але одночасно впливає й на процес виділення медіатора з пресинаптичних терміналей. Це досягається завдяки наявності в пресинаптичних закінченнях адренорецепторів. При взаємодії норадреналіну з пресинаптичним β-адренорецептором виділення медіатора щ знижується. І навпаки, при взаємодії норадреналіну з пресинаптичним β-адренорецептором виділення медіатора підсилюється. Норадреналін і адреналін при дії на гладкі м'язи активують розташовану в них аденілатциклазу. Вона, у свою чергу, при наявності іонів магнію, каталізує утворення цАМФ з АТФ, що активує енергетичний обмін. Норадреналін збільшує силу серцевих скорочень, викликає підвищення ж систолічного й діастолічного тиску без прискорення серцевого ритму. Унаслідок звуження ниркових судин норадреналін викликає гальмування діурезу й затримку натрію. Під впливом норадреналіну знижується секреція залоз шлунково-кишкового тракту, розслаблюються гладкі м'язи кишки, у к той же час слиновиділення підсилюється.
Адреналін характеризується більшим діапазоном симпатикотропної дії, ніж норадреналін. Збільшує хвилинний об'єм серця. Справляє розширюючу й антиспазматичну дію на бронхи, рефлекторно знижує частоту й амплітуду дихання. Адреналін є антидіуретиком, знижуючи виділення з сечею іоніь калію і натрію. Він гальмує шлунково-кишкову моторику, розслаблює стінки органів але скорочує сфінктери сечостатевої й шлунково-кишкової систем. гальмує травну секрецію. Адреналін підвищує скоротливість скелетних м'язів, що особливо виявляється на фоні їх втоми.