

Прокариоты (лат. Procaryota, от др.-греч. προ «перед» и κάρυον «ядро»), или доядерные — одноклеточные живые организмы, не обладающие (в отличие от эукариот) оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Для клеток прокариот характерно отсутствие ядерной оболочки, ДНКупакована без участия гистонов. Тип питания осмотрофный. Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), иархеи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды.
Прокариоты разделяют на два таксона в ранге домена (надцарства): Бактерии (Bacteria) и Археи (Archaea).[1]
Изучение бактерий привело к открытию горизонтального переноса генов, который был описан в Японии в 1959 г. Это процесс широко распространен среди прокариот, а также у некоторых эукариот. Открытие горизонтального переноса генов у прокариот заставило по-другому взглянуть на эволюцию жизни. Ранее эволюционная теория базировалась на том, что виды не могут обмениваться наследственной информацией. Прокариоты могут обмениваться генами между собой непосредственно (конъюгация, трансформация) а также с помощью вирусов — бактериофагов (трансдукция).
Изучением строения клетки занимается цитология (от латинского cytos – клетка и logos – учение).
Клетка – это ограниченная активной мембраной, упорядоченная, структурированная система биополимеров, образующих цитоплазму и ядро, участвующих в единой совокупности метаболических, энергетических и информационных процессов и осуществляющих поддержание и воспроизведение всей системы в целом (строение клетки изображено на рисунке 1.3.5). Это длинное и емкое определение требует дальнейших разъяснений.
Размер клеток может быть различным. Некоторые шаровидные бактерии имеют ничтожные размеры: от 0,2 до 0,5 мкм в диаметре (напомним, что 1 мкм в тысячу раз меньше 1 мм). В то же время существуют клетки, которые видны невооруженным глазом. Например, яйцо птицы – это, в сущности, одна клетка. Яйцо страуса достигает в длину 17,5 см, и это самая крупная клетка. Однако, как правило, размеры клеток колеблются в значительно более узких пределах – от 3 до 30 мкм.
Формы клеток также очень разнообразны. Некоторые из них приведены на рисунке 1.3.4. Клетки живых организмов могут иметь вид шара, многогранника, звезды, цилиндра и других фигур.
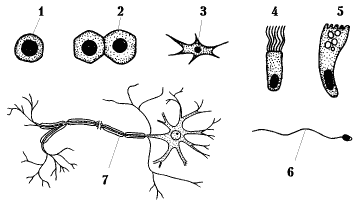
Рисунок 1.3.4. Формы клеток:
1 - клетка крови - лимфоцит; 2 - клетка печени - гепатоцит; 3 - клетка костной ткани - остеобласт; 4 - клетка мерцательного эпителия; 5 - бокаловидная клетка слизистой оболочки толстой кишки; 6 - мужская половая клетка (сперматозоид); 7 - клетка нервной ткани - нейрон
Несмотря на то, что клетки имеют разные формы и размеры, выполняют различные и часто весьма специфические функции, они, в принципе, имеют одинаковое строение, то есть у них можно выделить общие структурные единицы. Клетки животных и растений состоят из трех основных компонентов, представленных на рисунке 1.3.5: оболочки – клеточной мембраны (5), отделяющей содержимое клетки от внешней среды или от соседних клеток,цитоплазмы (4) и ядра (1).
|
Жгутиковые (Flagellata)
|
Это самый обширный класс простейших, к которому относится больше половины современных видов Protozoa. У жгутиковых тело шаровидное или удлиненное, с центрально расположенным ядром; на переднем конце тела имеется один или большее число (иногда много) тонких бичевидных жгутиков, с помощью которых животное передвигается. Некоторые жгутиковые захватывают пищу, выпуская псевдоподии, а другие, подобно ресничным инфузориям, имеют в определенном месте «рот» и «глотку». Большинство жгутиковых — очень мелкие организмы, и их трудно изучать, однако некоторые формы, например представители рода Euglena, достаточно крупны и в то же время обладают типичной для данного класса организацией. Эвглены содержат хлорофилл, находящийся в хлоропластах, и синтезируют значительную часть нужных им питательных веществ, но ни одна из них не является полным автотрофом. Здоровые культуры эвглен можно поддерживать только при добавлении к среде аминокислот. Тело эвглены покрыто нежной оболочкой — пелликулой, имеющей спиральные утолщения. Под пелликулой лежит слой сократимых нитей, что позволяет организму изменять свою форму. В цитоплазме разбросаны зеленые хлоропласты и прозрачные амилопласты, которые содержат резервные полисахариды. Единственный жгутик, основание которого находится в глубине глотки, образован путем слияния двух жгутиков. Эвглену, передвигающуюся с помощью одного жгутика, можно сравнить с человеком, который пытался бы плыть, действуя одной рукой. Жгутик при каждом своем ударе сгибается в сторону глазного пятна, но движется при этом не прямо назад, а косо по отношению к длинной оси тела. Поэтому тело не только отклоняется в сторону, но и слегка поворачивается вокруг своей длинной оси. Таким образом, последовательные удары жгутика заставляют эвглену продвигаться вперед по спирали, причем глазное пятно обращено кнаружи. Наибольшее число жгутиков и наиболее специализированное строение имеют жгутиковые, обитающие в кишечнике термитов. Некоторые представители этого класса обладают признаками, свойственными другим классам простейших или даже губкам; эти формы указывают на некоторые промежуточные этапы тех путей, по которым, возможно, шло развитие других простейших и губок от жгутиковых. Представители жгутиковых, более сходные с растениями, например колониальное жгутиковое Volvox. Клетки, образующие колонию Volvox, соединены между собой цитоплазматическими мостиками, через которые может осуществляться синхронизация активности различных клеток. Виды, более близкие к настоящим животным, мелки и не так широко распространены. Особый интерес представляет группа Choanoflagellata, представители которой сходны с воротничковыми клетками губок. Это сидячие формы, прикрепленные стебельком к субстрату, а их единственный жгутик окружен у основания тонким «воротничком» из цитоплазмы. Среди жгутиковых есть и паразиты, вызывающие заболевания человека; таковы, например, трипаносомы, вызывающие сонную болезнь и передаваемые от человека к человеку мухой це-це. Жгутиковых обычно рассматривают как исходную группу, от которой в процессе эволюции произошли не только другие группы простейших, но также высшие растения и животные.
|
Для эволюции прокариотных организмов характерен ярко выраженный физиолого-биохимический уклон, т.е. основное развитие прокариот шло по линии формирования и опробования различных функций, результатом чего и явилось сегодняшнее многообразие типов жизни в микромире. Поразительное физиологическое разнообразие прокариот сформировано на базе весьма ограниченного числа морфологических форм. Действительно, морфологическая эволюция прокариот прошла незначительный путь, так что мы можем говорить лишь о зачатках морфологической дифференцировки на базе прокариотной клеточной организации. Относительно "продвинутыми" в этом направлении оказались толькоэубактерий. Дляархебактерийхарактерно отсутствие сложных морфологических форм и какой-либо клеточной дифференцировки. Вегетативные клетки многих эубактерий в определенных условиях дают начало структурам, морфологически отличающимся от исходных. Ими могут быть вегетативные клетки, но измененной формы, клеточные структуры с четко выраженной функциональной специализацией, различные многоклеточные образования. В подавляющем большинстве случаев все известные проявления морфологической дифференцировки эубактерий направлены на повышение их выживаемости. Это выражается как в формировании специальных клеток, обладающих повышенной устойчивостью к перенесению неблагоприятных условий ( эндоспоры,цисты), так и в формировании структур, обеспечивающих эффективное размножение вида ( гормогонииибаеоцитыцианобактерий). В основе морфологической дифференцировки лежат определенные биохимические процессы, которые, в свою очередь, являются выражением соответствующей генетической информации. (Ф.Жакоб (F.Jacob) и Ж.Моно (J.Monod) определили дифференцировку следующим образом: "Одну клетку следует считать дифференцированной по сравнению с другой, если при одинаковых геномах набор белков, синтезированых в этих клетках, различен"). Генетическая информация запрограммирована в генетическом аппарате клетки и реализуется в процессе ее развития или же в зависимости от действия различных внешних факторов. Образование морфологически дифференцированных клеток у разных представителей эубактерий суммировано втабл. 6. Большинство таких структур относится к категории покоящихся форм, назначение которых - обеспечить переживание вида в течение длительного времени в неблагоприятных условиях. Этоэндоспорырядаграмположительных бактерий,цистыазотобактераимиксобактерий,акинетыцианобактерий, экзоспоры отдельных представителейметилотрофных бактерийифототрофных бактерий, экзо- и эндоспорыактиномицетов. После попадания в подходящие условия покоящиеся формы прорастают, давая начало вегетативным клеткам. Другие морфологически дифференцированные клетки служат для размножения. К ним относятся, например,гормогонииибаеоцитыцианобактерий. Наконец, третьи ( гетероцистыцианобактерий,бактероидыклубеньковых бактерий) связаны с осуществлением уникального процесса, свойственного только прокариотным организмам, - фиксацией молекулярного азота атмосферы. |
Классификация водорослей
Разделение водорослей на систематические группы (таксоны) высшего ранга — отделы (Divisio, Phylum) — в основном совпадает с характером их окраски, связанной, конечно, с особенностями строения. Однако относительно количества и объема этих отделов в научной литературе нет единства взглядов. В настоящем издании мы принимаем классификацию, наиболее широко распространенную в научной литературе, а именно деление водорослей на 10 отделов. Приводим их перечень с соблюдением общепринятой русской и латинской научной номенклатуры:
1) сине-зеленые водоросли — Cyanophyta; 2) пирофитовые водоросли — Pyrrophyta; 3) золотистые водоросли — Chrysophyta; 4) диатомовые водоросли — Bacillariophyta; 5) желто-зеленые водоросли — Xanthophyta; 6) бурые водоросли — Phaeophyta; 7) красные водоросли — Rhodophyta; 8) эвгленовые водоросли — Euglenophyta; 9) зеленые водоросли — Chlorophyta; 10) харовые водоросли — Charophyta.
В научной литературе до сих пор продолжа- ются споры о положении в общей системе, с одной стороны, сине-зеленых водорослей и, с другой стороны, всех тех водорослей, которые представлены одноклеточными подвижными формами, снабженными органами движения — жгутиками (это почти все эвгленовые водоросли, большая часть пирофитовых и золотистых во- дорослей и отдельные классы желто-зеленых и зеленых водорослей).
Действительно, сине-зеленые водоросли резко отличаются от других водорослей простотой внутренней организации клеток. Клетки их лишены оформленного ядра, что сближает их с бактериями. Вместе с бактериями сине-зеленые водоросли составляют раздел организмов, обозначаемый как прокариоты (Prokaryota), т. е. «доядерные», в отличие от всех остальных растений и животных, обладаю- щих оформленным клеточным ядром и обозначаемых как эукариоты (Eukaryota), т. е. «истинно ядерные». Некоторые авторы придают этому признаку основное таксономическое значение и объединяют сине-зеленые водоросли с бактериями, т. е. вообще исключают их из числа собственно водорослей.
Что же касается жгутиковых форм водорослей, то здесь вопрос осложняется тем, что они во многих случаях близки к подобным же бесцветным формам, что дало повод для объединения всех их в общую систематическую груп- пу «жгутиковых организмов» и включения в систему животного мира. Такая тенденция сохранилась и до сих пор. В современной зоологической систематике жгутиковые рассмат- риваются как один из классов — жгутиконосцы (Mastigophora, или Flagellata) типа одноклеточных животных — простейших (Protozoa), а в пределах этого класса окрашенные жгутиковые объединяются в подкласс растительные жгутиконосцы (Phytomastigina), бесцветные — в подкласс животные жгутиконосцы (Zoomastigina).
Не вдаваясь здесь в разбор как первого, так и второго вопроса (это уместнее сделать после описания всех водорослей), отметим только, что мы не можем принять указанные крайние точки зрения. Дело в том, что в процессе эволюции возникновение хлорофилла у первичных бесцветных гетеротрофных организмов было настолько существенным переломным моментом, что только его и можно считать началом той богатейшей эволюции, которая привела к созданию биосферы на Земле в ее современном выражении. И трудно допустить, что такое сложное и уникальное органическое соединение, как хлорофилл, давшее организмам возможность фототрофного питания, т. е. совершенно новой энергетики жизненных процессов, основанной на утилизации солнечной энергии, могло возникать многократно на разных этапах эволюции. С этих позиций мир водорослей как первичных фототрофных организмов един и целостен. Морфологическое многообразие его различных ветвей есть следствие эволюционного взрыва, вызванного появлением фотосинтеза, который обеспечил хлорофиллоносным организмам успешное развитие в чисто абиотической среде. Учитывая особенности строения клеток сине-зеленых водорослей, следует думать, что возникновение хлорофилла произошло еще на прокариотическом уровне, а наличие в настоящее время сходных хлорофиллоносных и бесцветных эукариотических жгутиковых форм обусловлено морфологическим параллелизмом эволюционного развития в разных ветвях организмов. Во всяком случае, у водорослей подобное явление морфологического параллелизма распространено очень широко (например, строение в пределах Chlorophyta, Xanthophyta и Chrysophyta). Такая точка зрения хорошо подтверждается еще и тем, что в пределах большинства вышеперечисленных отделов водорослей жгутиковые формы тесно связаны переходами с другими, типично «водорослевыми» структурами — неподвижными клетками, колониями и нитями. С другой стороны, в пределах некоторых отделов имеются и безусловно вторично обесцветившиеся формы.
Таким образом, у нас нет оснований отказываться от рассмотрения водорослей как морфо-физиологической целостности, от выяснения их многообразия в целом, происхождения и взаимных филогенетических связей. Точно так же с этих позиций целостности хорошо выявляются место и роль водорослей в природе: в историческом плане они представляют собой первый этап в развитии всего зеленого ствола растительного мира, а в общем круговороте веществ в природе играют огромную роль как первичное звено всех пищевых связей в водной среде и гигантский поставщик кислорода в атмосферу.
Изучение всех этих вопросов составляет предмет особой науки — альгологии (от латинского названия водорослей — algae).