

32. Артериальные (периферические) и центральные хеморецепторы, их роль в регуляции вентиляции легких.
Артериальные
(периферические) хеморецепто- р ы.
Периферические хеморецепторы находятся
в каротидных и аортальных тельцах.
Сигналы от артериальных хеморецепторов
по синокаротидным и аортальным нервам
первоначально поступают к нейронам
ядра одиночного пучка продолговатого
мозга, а затем переключаются на нейроны
дыхательного центра. Ответ периферических
хеморецепторов на понижение Рао2 является
очень быстрым, но нелинейным. При Рао2
в пределах 80—60 мм рт. ст. (10,6—8,0 кПа)
наблюдается слабое усиление вентиляции,
а при Рао2 ниже 50 мм рт. ст. (6,7 кПа) возникает
выраженная гипервентиляция.
Расог
и pH крови только потенцируют эффект
гипоксии на артериальные хеморецепторы
и не являются адекватными раздражителями
для этого типа хеморецепторов
дыхания.
Реакция
артериальных хеморецепторов и дыхания
на гипоксию. Недостаток 02 в артериальной
крови является основным раздражителем
периферических хеморецепторов. Импульсная
активность в афферентных волокнах
синокаротидного нерва прекращается
при Расgt;2 выше 400 мм рт. ст. (53,2 кПа). При
нормоксии частота разрядов синокаротидного
нерва составляет 10% от их максимальной
реакции, которая наблюдается при Рао2
около 50 мм рт. ст. и ниже. Гипоксическая
реакция дыхания практически отсутствует
у коренных жителей высокогорья и исчезает
примерно через 5 лет у жителей равнин
после начала их апаптации к высокогорью
(3500 м и выше).
Центральные
хеморецепторы. Окончательно не установлено
местоположение центральных хеморецепторов.
Исследователи считают, что такие
хеморецепторы находятся в ростральных
отделах продолговатого мозга вблизи
его вентральной поверхности, а также в
различных зонах дорсального дыхательного
ядра.
Наличие
центральных хеморецепторов доказывается
достаточно просто: после перерезки
синокаротидных и аортальных нервов у
подопытных животных исчезает
чувствительность дыхательного центра
к гипоксии, но полностью сохраняется
реакция дыхания на гиперкапнию и ацидоз.
Перерезка ствола мозга непосредственно
выше продолговатого мозга не влияет на
характер этой реакции.
Адекватным
раздражителем для центральных
хеморецепторов является изменение
концентрации Н* во внеклеточной жидкости
мозга. Функцию регулятора пороговых
сдвигов pH в области центральных
хеморецепторов выполняют структуры
гематоэнцефалического барьера, который
отделяет кровь от внеклеточной жидкости
мозга. Через этот барьер осуществляется
транспорт 02, С02 и Н+ между кровью и
внеклеточной жидкостью мозга. Транспорт
С02 и Н+ из внутренней среды мозга в плазму
крови через структуры гематоэнцефалического
барьера регулируется с участием фермента
карбоангидразы.
Реакция
дыхания на С02. Гиперкапния и ацидоз
стимулируют, а гипокапния и алкалоз
тормозят центральные хеморецепторы.
Для
определения чувствительности центральных
хеморецепторов к изменению pH внеклеточной
жидкости мозга используют метод
возвратного дыхания. Испытуемый дышит
из замкнутой емкости, заполненной
предварительно чистым 02. При дыхании в
замкнутой
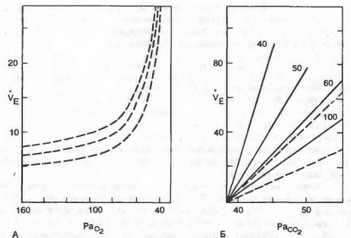 Рис.
8.12. Изменение вентиляции легких (Ve, л*мии
1) в зависимости от парциального давления
Ог (А) и СО2 (Б) в альвеолярном воздухе
при различном содержании Ог в альвеолярном
воздухе (40, 50, 60 и 100 мм рт. ст.).
В
системе
выдыхаемый СО2 вызывает линейное
увеличение концентрации СО2 и одновременно
повышает концентрацию ЬГ в крови, а
также во внеклеточной жидкости мозга.
Тест проводят в течение
Рис.
8.12. Изменение вентиляции легких (Ve, л*мии
1) в зависимости от парциального давления
Ог (А) и СО2 (Б) в альвеолярном воздухе
при различном содержании Ог в альвеолярном
воздухе (40, 50, 60 и 100 мм рт. ст.).
В
системе
выдыхаемый СО2 вызывает линейное
увеличение концентрации СО2 и одновременно
повышает концентрацию ЬГ в крови, а
также во внеклеточной жидкости мозга.
Тест проводят в течение
5 мин под контролем содержания СО2 в выдыхаемом воздухе.
На рис. 8.12 показано изменение объема вентиляции при различном уровне напряжения СОг в артериальной крови. При Расог ниже 40 мм рт. ст. (5,3 кПа) может возникнуть апноэ в результате гипокапнии. В этот период дыхательный центр мало чувствителен к гипоксической стимуляции периферических хеморецепторов.