

2 курс / Нормальная физиология / Экзамен Физиология / tema_1_fiziologia_sinapsov
.docx

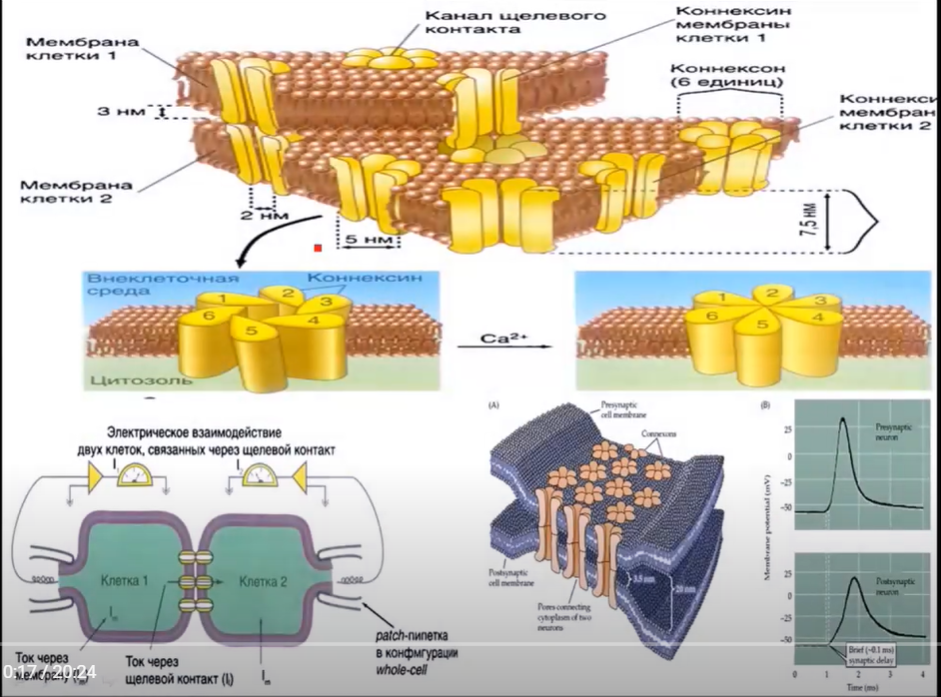
Проведение возбуждения в электрических синапсах обусловлено узкой синаптической щелью и наличием пре- и постсинаптической мембраны, интегральных белков, коннексонов. Эти белки образованы из нескольких субъединиц, которые называются коннексины. Внутри них есть просвет, так что каждая пара расположенных по одной линии коннексонов образует канал, через который сообщаются 2 клетки. Просвет этих каналов контролируется ионами кальция, а также протонами водорода. При повышении концентрации кальция и накоплении протонов водорода, т.е. снижение рН и ацидоз, происходит закрытие этих каналов. Благодаря узкой синаптической щели возникший потенциал действия на пресинаптической мембране электротонически за счет локальных токов передается на постсинаптическую мембрану, а через белки-коннексоны натриевый ток, проходящий внутрь пресинаптической мембраны поступает в постсинаптическую мембрану, вызывая там местную деполяризацию.
В организме между нейронами человека электрические синапсы мало распространены. Однако, такие щелевые контакты или нексусы распространены между кардиомиоцитами, а также между ГМК. Такие щелевые контакты обеспечивают синхронность возбуждения и сокращения.
Нарушение обмена веществ в кардиомиоцитах, например при снижении кровоснабжения миокарда, сопровождается повышением протонов водорода, т.е. ацидозом, что приводит к закрытию каналов-коннексонов и как следствие к разобщению кардиомиоцитов, например при инфаркте миокарда.

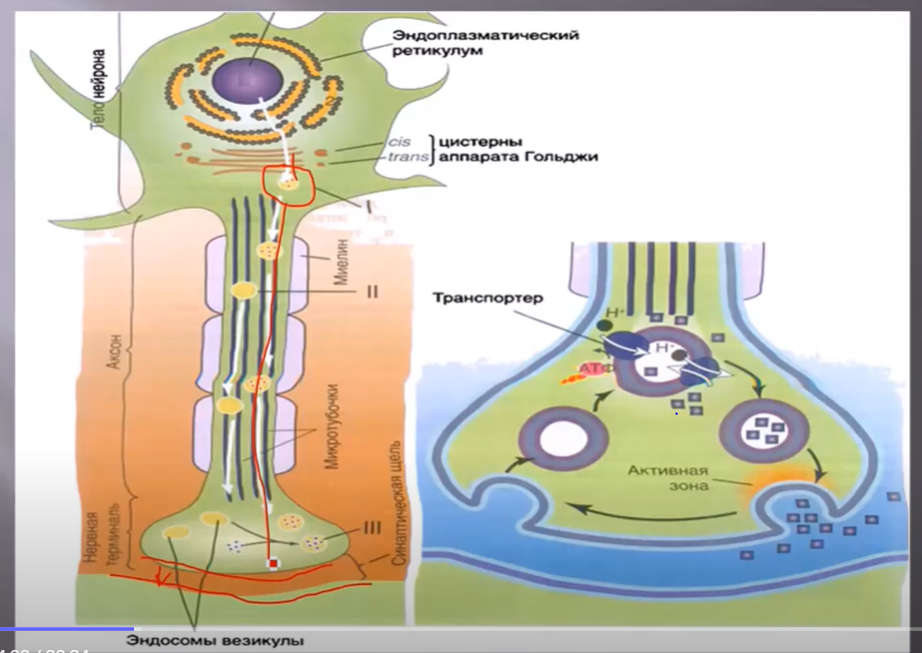
Химические синапсы, также как и электрические, состоят из пресинаптической мембраны, синаптической щели и постсинаптическоц мембраны. Однако синаптическая щель здесь шире и передача информации происходит за счет выделения медиатора или нейротрансмиттера. Везикулы и предшественники пептидных нейротрансмиттеров высвобождаются из аппарата Гольджи и путем аксонального транспорта доставляются до терминали аксона, т.е. ближе к пресинаптической мембране. При этом пептидные трансмиттеры уже заключены в везикулы.
Непептидные медиаторы, например норадреналин, ацетилхолин, серотонин, дофамин, тормозные медиаторы (ГАМК и глицин) синтезируются и транспортируются в везикулы в нервных окончаниях, с образованием лабильных и стабильных фракций этого медиатора. Процесс упаковки медиатора в везикулу происходит за счет электронно-протонного насоса с затратой энергии АТФ.
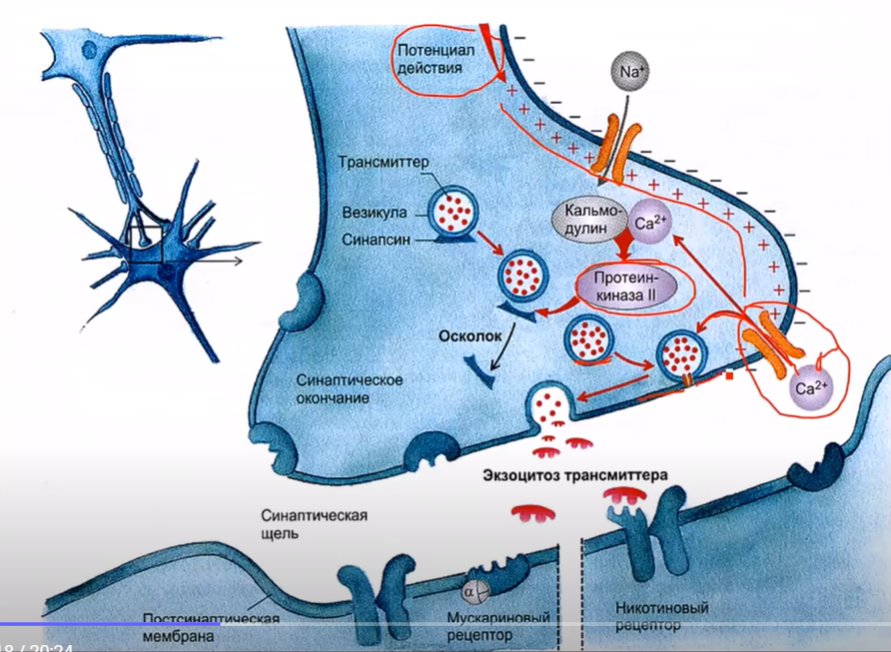
Процесс выделения медиатора начинается достижением потенциала действия до терминали аксона. Вследствие этого открываются потенциал-управляемые кальциевые каналы. Кальций из синаптической щели путем диффузии, т.е. по градиенту концентрации поступает в пресинаптическую мембрану, при этом активируя внутриклеточные ферменты протеинкиназы, что в свою очередь способствует активации специализированных белков, обеспечивающих слияние мембраны везикулы с пресинаптической мембраной. В результате этого возникает экзоцитоз с последующим выделением медиатора.
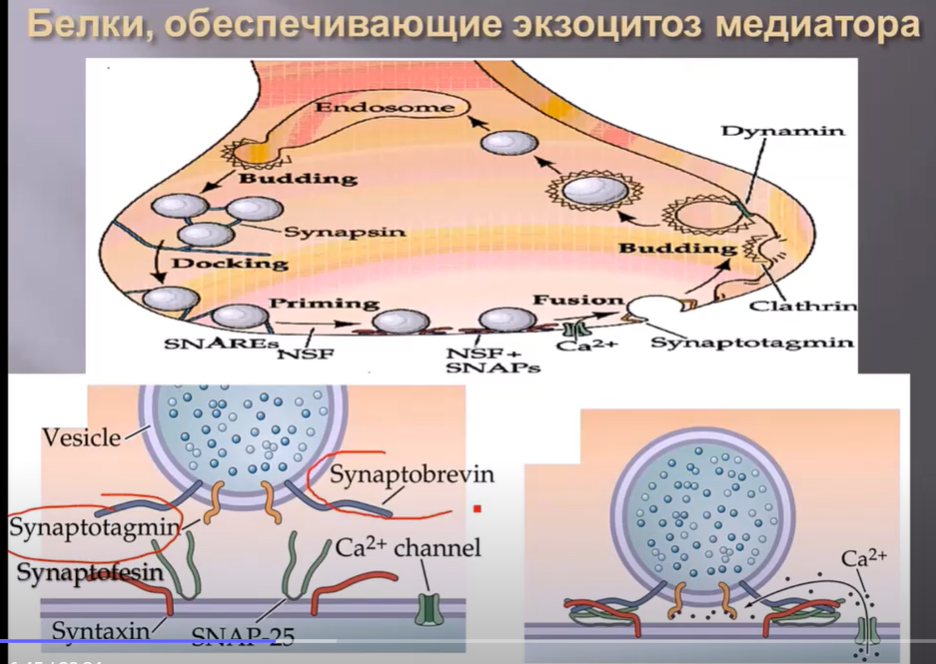
Белки, участвующие в экзоцитозе медиатора, расположены как в мембране везикулы, например такие как синаптотагмин, синаптобревин, так и в пресинаптической мембране, такие как синтаксин и снэп-25. Но только благодаря входу кальция в пресинаптическую мембрану, и в частности взаимодействию с синаптотагмином, запускается процесс слияния везикулы с пресинаптичской мембраной, благодаря белковым структурам.
Повреждение этих белков естественно сопровождается нарушением экзоцитоза. Например, ботулотоксин разрушает синаптобревин, синтаксин и снэп-25. Это происходит в нервно-мышечных синапсах, что сопровождается снижением выделения медиатора ацетилхолина и как следствие расслабление мышц.
Столбняный токсин разрушает в основном белок синаптобревин в тормозных синапсах, что приводит к развитию ригидности мышц.
Латротоксин, продуцируемый пауком черная вдова, нарушает функцию синаптотагмина.
Тем не менее, некоторые из этих токсинов используются в медицине, в частности ботулотоксин. Впервые в медицинской практике в конце 70-хх годов ботулотоксин был использован офтальмологом Алан Скотом для лечения блефароспазма. В современной медицине этот токсин используется для снятия ригидности мышц, а в косметологии для разглаживания мимических морщин.
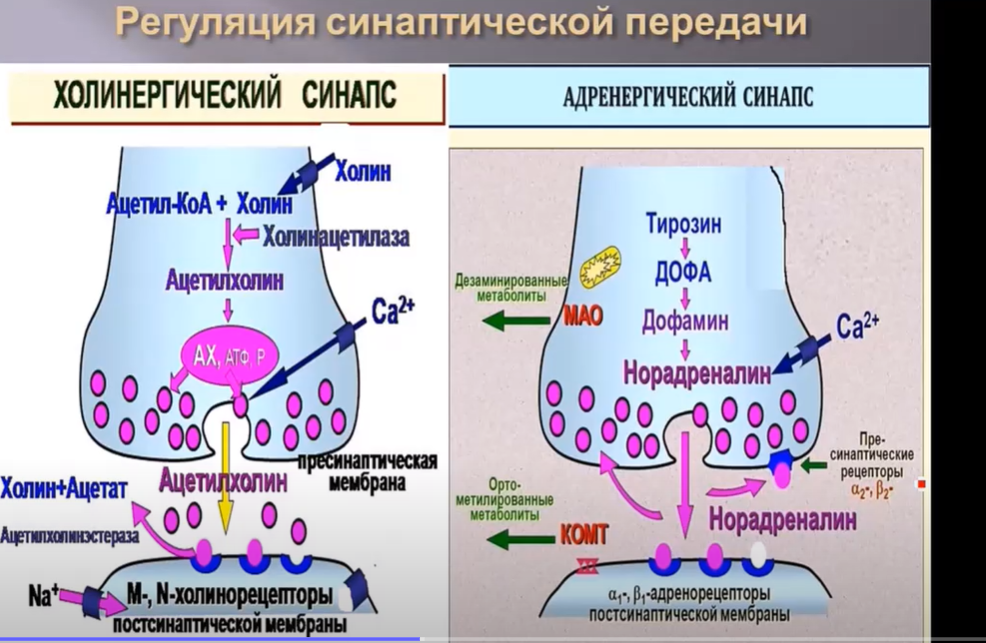
В естественных условиях в организме человека регуляция синаптической передачи происходит как на уровне синтеза самого медиатора, так и на уровне аксонального транспорта, а также на уровне пресинаптической мембраны, синаптической щели и постсинаптической мембраны. В частности, на уровне пресинаптической мембраны регуляция выделения медиатора осуществляется несколькими путями. Например, в синапсах осуществляется обратный нейральный захват, т.е. реаптейк избытка медиатора. Кроме того, в пресинаптической мембране существуют пресинаптические рецепторы, в адренергических синапсах это альфа2 и бетта2 адренорецепторы. При взаимодействии с этими рецепторами последующее выделение медиатора изменяется, а именно при взаимодействии с альфа2 адренорецепторами последующая секреция медиатора уменьшается, а при взаимодействии с бетта-адренорецепторами увеличивается. По такому же принципу контролируетс регуляция выделения ацетилхолина, в частности при взаимодействии ацетилхолина на пресинаптической мембране с M-холинорецепторами последующее выделение медиатора уменьшается, а при взаимодействии с Н-никотинчувствительными рецепторами последующее выделение медиатора увеличивается.
В адренергических синапсах на уровне пресинаптической мембраны работает фермент моноаминоксидаза. Этот фермент в терминали аксона разрушает, а точнее обеспечивает дезаминирование и снижение активности норадреналина. В синаптической щели работают эндогенные регуляторы.
В холинергических синапсах работает фермент ацетилхолинэстераза. Данный фермент разрушает ацетилхолин и тем самым препятствует избыточному действию медиатора.
В синаптической щели адренергических синапсов работает другой фермент катехоламинометилтрансфераза и за счет метилирования снижает активность норадреналина.
В свою очередь в медицинской практике широко используются препараты, влияющие на синаптическую передачу в химических синапсах. Одни из них контролируют механизмы обратного нейрального захвата, другие влияют на активность ферментов. Например, ингибиторы моноаминоксидазы повышают концентрацию норадреналина между нейронами структур лимбической системы и тем самым относятся к антидепрессантам. Ингибиторы ацетилхолинэстеразы в нервно-мышечных синапсах используются при лечении миастении и наоборот, реактиваторы ацтилхолинэстеразы используются при отравлении фосфоорганическими соединениями.
Кроме того, существует огромное количество лекарственных средств, которые влияют на рецепторы пре- и постсинаптической мембраны, активируя или блокируя их и тем самым оказывая эффект миметиков или блокаторов. Например, хлороподобные вещества в нервно-мышечном синапсе блокируют Н-холинорецепторы и тем самым расслабляют мышцу, вызывая эффект миорелаксации.
Выделившиеся медиаторы в синаптическую щель взаимодействуют с рецепторами на постсинаптической мембране.
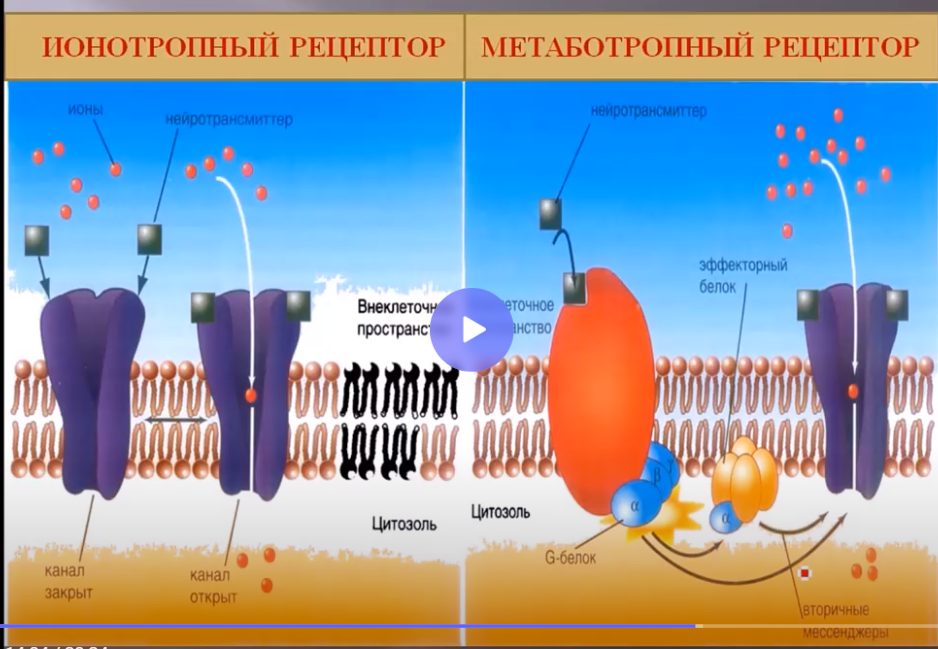
В зависимости от того, каким путем активируются каналы на постсинаптической мембране, существуют ионотропные рецепторы и метаботропные рецепторы. Эти рецепторы изменяют активность ионных каналов разными механизмами. В частности, ионотропный рецептор, например никотинчувствительные рецепторы в скелетных мышцах или в вегетативном ганглии, при этом Н1-рецепторы расположены в нервно-мышечных синапсах, а Н2 на постсинаптической мембране нейронов вегетативного ганглия. Активация таких ионотропных рецепторов приводит непосредственно к открытию ионных каналов, в частности натриевых каналов, в результате чего возникает местный постсинаптический процесс. Если же это метаботропный рецептор, то открытие ионных каналов проходит через систему вторичных посредников, например через активацию G-белка и вторичных посредников –цАМФ, цГМФ. К таким метаботропным рецепторам относится М-холинорецепторы в кардиомиоцитах.
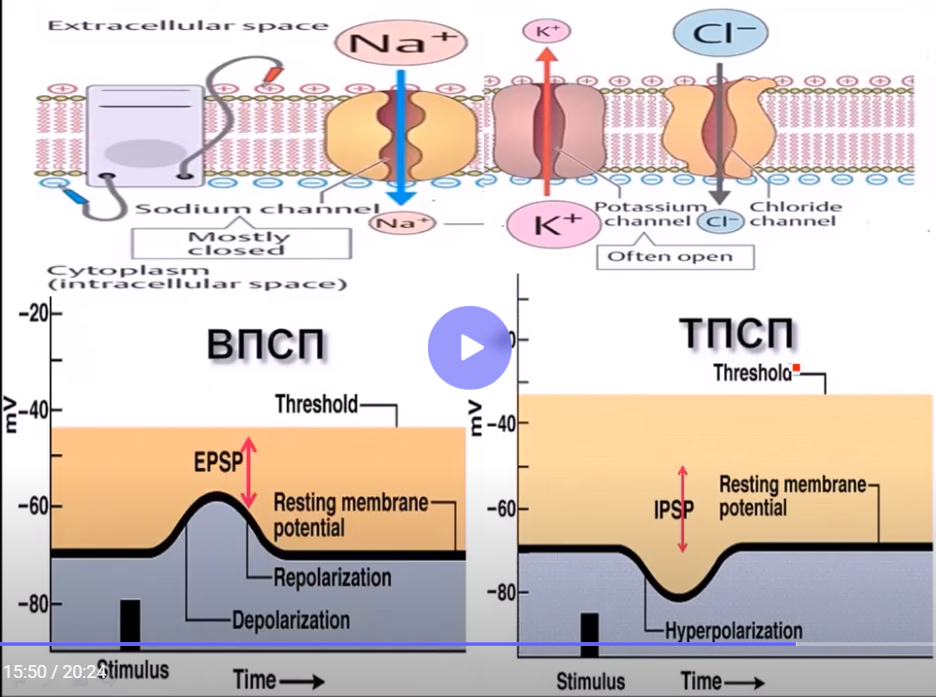
В нервных синапсах взаимодействие медиатора с рецепторами приводит к изменению активности ионных каналов на постсинаптической мембране. И в зависимости от того, какой это медиатор, т.е. возбуждающий или тормозящий, на постсинаптической мембране соответственно возникает местный процесс в виде ВПСП или ТПСП. Возбуждающий медиатор – норадренали или ацетилхолин – на постсинаптической мембране нервной клетки вызывает открытие натриевых каналов из m-ворот. За счет диффузии натрий поступает в клетку с развитием местной деполяризации, при этом происходит приближение к критическому уровню и тем самым возбудимость увеличивается за счет снижения порога раздражения. Если же это медиатор тормозный – глицин или ГАМК – то эти медиаторы влияют на калиевые и хлорные каналы, а именно усиливают выход калия из клетки и вход хлора в клетку, тем самым увеличивая уровень отрицательности на постсинаптической мембране с развитием гиперполяризации. В результате этого происходит увеличение порога раздражения и как следствие снижение возбудимости и тем самым возникает торможение нервной клетки.



Сеченовское (центральное) торможение

