

2 курс / Гистология / эндокринная
.pdfГЛАВА 16
ЭНДОКРИННАЯ СИСТЕМА
ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА ЭНДОКРИННОЙ СИСТЕМЫ. ПОНЯТИЕ ОБ АУТОКРИНИИ, ПА-
РАКРИНИИ И ЭНДОКРИНИИ
Эндокринная система относится к числу регуляторно-интегрирующих систем организма наряду с нервной и иммунной системами, находясь с ни-
ми в теснейшем взаимодействии. К ее функциям относится регуляция
важнейших вегетативных функций организма: соматического роста, репродукции; размножения, роста и дифференцировки клеток; обмена веществ и энергии; секреции; экскреции; всасывания; сокращения гладкой и сердечной мышечных тканей; поведенческих реакций; процессы адаптации и др. В целом функции эндокринной системы можно определить как поддержание гомеостаза организма при помощи секретируемых во внутреннюю среду организма гормонов (гуморальная регуляция гомеостаза).
Гуморальная регуляция гомеостаза в процессе эволюции претерпела существенные изменения. Это нашло отражение в виде формирования нескольких ее уровней, которые существуют у современных животных и чело-
века (Рис. 16.1).
1.Аутокриния. В этом случае секретируемые клеткой гормоны (аутокоиды) регулируют собственную функциональную активность или функциональную активность клетки этого же типа, регулируя их активность. В качестве одного из примеров можно привести кейлоны, которые вырабатываются дифференцированными клетками дифферона и влияют на малодифференцированные клетки этого дифферона, подавляя их митотическую активность. Например, продуцируемые более дифференцированными кератиноцитами эпидермальные кейлоны, подавляя митотическую активность базальных кератиноцитов, регулируют тканевой гомеостаз в эпидермисе. В- лимфоциты вырабатывают медиаторы, стимулирующие их собственную дифференцировку и превращение в плазмоциты. Нейтрофильные лейкоциты выделяют вещества, которые воздействуют на другие нейтрофильные лейкоциты и регулируют их активность.
2.Паракриния. При данном виде гуморальной регуляции выделяемые клеткой одного вида биологически активные вещества (гормоны и гормоноподобные вещества) действуют на активность рядом расположенных клеток другого типа. Например, входящие в состав желудочных желез G-клетки секретируют гормон гастрин, изменяющий функции главных и парие-
129
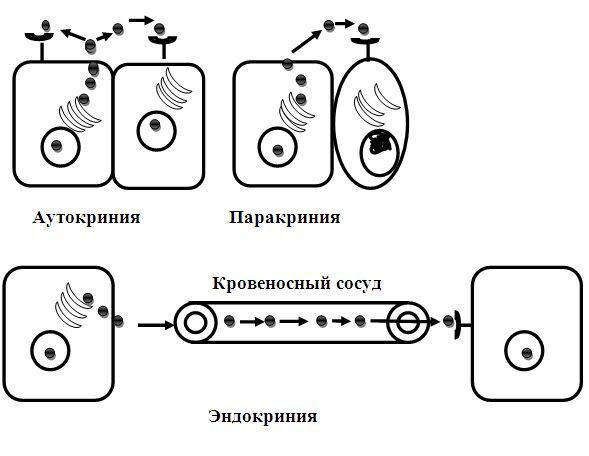
тальных клеток, входящих в состав этих же желез и продуцирующих соответственно пепсин и соляную кислоту.
Рис. 16.1. Варианты действия биологически активных веществ (гормонов, аутокоидов и др.): при аутокринии продуцируемые клеткой молекулы биологически активные веществ (БАВ) действуют на собственные рецепторы и на рецепторы клеток этого же типа; паракриния характеризуется действием БАВ на соседние, но другого типа клетки этой же ткани; при эндокринии секреция гормона происходит во внутреннюю среду, затем в сосуды (кровеносные, лимфатические) и с током крови или лимфы достигают клетки-мишени, действуют на ее рецепторы и изменяя ее функциональное состояние
3 Эндокриния - такой способ гуморальной регуляции, при котором вырабатываемые клеткой регуляторные факторы удалены от регулируемых ими клеток на достаточно большое расстояние. В этом случае в качестве примера можно рассматривать любую эндокринную железу.
СОСТАВ ЭНДОКРИННОЙ СИСТЕМЫ
Эндокринная система состоит из: 1) органов - эндокринных желез, вырабатывающих гормоны (щитовидная железа, надпочечник, эпифиз, ги-
пофиз и др.). 2) эндокринных частей неэндокринных органов (островки
130
Лангерганса поджелудочной железы и др.). 3) одиночных гормонпродуцирующих клеток, расположенных диффузно в различных органах. Совокупность таких клеток образует так называемую диффузную эндокринную
систему (ДЭС).
ОБЩИЕ ПРИНЦИПЫ СТРУКТУРНО-ФУНКЦИОНАЛЬНОЙ ОРГАНИЗАЦИИ ЭНДОКРИННЫХ ЖЕЛЕЗ
Несмотря на различия в строении, все эндокринные железы имеют общие морфофункциональные признаки.
1.Эти железы не имеют выводных протоков, т.к. выделяют секреты (гормоны) непосредственно в кровь, лимфу или окружающую клетки тканевую жидкость.
2.В связи с этим они богато кровоснабжаются.
3.Т.к. секрет эндокринных желез должен быстро достигать регулируемых органов, тканей или клеток, для облегчения выхода гормонов в кровь капилляры эндокринных желез являются капиллярами или фенестрированного, или синусоидного типа.
4.Эндокринные железы относятся к органам паренхиматозного типа и
вбольшинстве случаев образованы эпителиальной тканью, формирующей тяжи или фолликулы. Наряду с этим, в ряде эндокринных желез и ткани других типов могут являться функционально ведущими. Так, например, в гипоталамусе и эпифизе таковой является нервная ткань. Юкстагломерулярные клетки почки, вырабатывающие ренин, и эндокринные кардиомиоциты миокарда, продуцирующие натрийуретический фактор, относятся к мышечным тканям, а интерстициальные клетки почек и гонад, адипоциты жировой ткани являются соединительнотканными клетками.
5.В эндокринных органах преобладает паренхима, строма же развита слабее, т.е. эти органы построены экономно.
6.Секреты эндокринных органов (гормоны, гормоноиды и другие биологически активные вещества) оказывают на мишени выраженные эффекты в малых количествах.
7.Имеются некоторые общие принципы строения гормонпродуцирующих клеток, входящих в состав эндокринных образований.
КЛАССИФИКАЦИЯ ГОРМОНОВ. Строение эндокриноцитов, входящих в состав эндокринных желез, во многом зависит от химического строения гормона. Все вырабатываемые эндокринными железами и клетками гормоны можно разделить на три группы.
1. Гормоны белковой, гликопротеиновой и полипептидной природы.
Таковыми являются гормоны гипофиза, гипоталамуса, поджелудочной и
131
некоторых других желез.
2.Гормоны - производные аминокислот. К ним относятся гормоны щитовидной железы тироксин и трийодтиронин, гормон мозгового вещества надпочечников адреналин, серотонин, мелатонин, вырабатываемый многими эндокринными железами и клетками, и др.
3.Стероидные гормоны. Это гормоны-производные холестерола. К стероидным гормонам относятся половые гормоны, гормоны коры надпочечников, витамин D3 (кальцитриол).
Вдальнейшем будет продемонстрирована зависимость строения эндокринных клеток, составляющих эндокринную железу, и их секреторного цикла от химической природы вырабатываемых гормонов.
МЕХАНИЗМ ДЕЙСТВИЯ ГОРМОНОВ. Попадая в кровь (лимфу, тканевую жидкость), гормоны с ее током достигают регулируемых клеток, тканей, органов, которые называются мишенями. Таким образом, молекулы гормона являются первичными посредниками (мессенджерами) между эндокринной железой и ее мишенью. Можно выделить два основных механизма действия гормонов:
1.Гормон представляет собой полярную молекулу, которая не может проникнуть в клетку через ее оболочку. В этом случае гормон взаимодействует с поверхностными рецепторами клеток и изменяет пространственную конфигурацию этих рецепторов. Рецепторы являются трансмембранными белками и, как правило, относятся либо к рецепторам, связанным с G- белками, либо к каталитическим рецепторам. G-белки активируют или ингибируют ряд внутриклеточных эффекторов, в том числе и ферменты, выра-
батывающие вторичные, внутриклеточные посредники (циклический АМФ, циклический ГМФ, инозитолтрифосфат, диацилглицерол и др.) и изменяющие внутриклеточный метаболизм. Такими ферментами, например, являются фосфодиэстераза, фосфолипазы, аденилатциклаза.
Каталитические рецепторы состоят из поверхностной рецепторной, трансмембранной и внутренней каталитической частей. При связывании гормона с таким рецептором активируется каталитическая субъединица, запускающая в клетке каскад биохимических реакций. Примером такого трансмембранного каталитического рецептора-фермента является тирозинкиназа, включающая в клетке фосфорилирование компонентов так называемого ras-каскада и как итог - транскрипцию генов, обеспечивающих пролиферацию. Через каталитические рецепторы свое действие на клетку оказывают инсулин, факторы роста и некоторые другие полипептидные гормоны. Мутации генов ras-каскада приводят к злокачественному росту клеток.
2.При реализации второго механизма гормон вследствие своей непо-
132
лярности и липофильности проникает в клетку, связывается с внутриклеточным белком-рецептором и вместе с ним транспортируется в ядро клетки, где изменяет активность соответствующих генов. Это ведет к изменению метаболизма клетки. Для таких гормонов в ядре клеток-мишеней существуют ядерные рецепторы. Они состоят из двух частей: 1) участка, связывающего гормон, и 2) участка, являющегося фактором транскрипции и взаимодействующего со специфическими последовательностями ДНК клетки-мишени. Эти же гормоны могут действовать на органеллы клетки. Например, митохондрии являются мишенями для тиреоидных гормонов. В таком случае рецепторы к гормону имеются и на мембранах органелл. Ко второй группе гормонов относятся жирорастворимые неполярные стероидные, тиреоидные гормоны, витамин D3, которые благодаря липотропным свойствам легко проникают внутрь клетки через ее оболочку.
КЛАССИФИКАЦИЯ ЭНДОКРИННЫХ ЖЕЛЕЗ
I.КЛАССИФИКАЦИЯ ПО ИЕРАРХИЧЕСКОМУ ПРИНЦИПУ
1.Центральные эндокринные органы. К этим органам относятся ги-
поталамус, эпифиз и гипофиз. Они переключают нервные влияния на эндокринные, осуществляют контроль за деятельностью других (периферических) эндокринных желез.
2.Периферические эндокринные органы. Эти эндокринные железы осуществляют непосредственный контроль над важнейшими функциями организма. В зависимости от того, находятся или не находятся они под регулирующим влиянием гипофиза, периферические эндокринные железы делятся на две группы:
2.1.Аденогипофизнезависимые периферические эндокринные железы и эндокринные клетки. К ним относятся: С-тироциты (парафолликулярные клетки) щитовидной железы, паращитовидные железы, мозговое вещество надпочечников, островковый аппарат поджелудочной железы, ретикулоэпителиоциты тимуса, клубочковая зона коры надпочечника, большинство эндокринных клеток ДЭС.
2.2.Периферические эндокринные железы, зависимые от аденоги-
пофиза. Регулирующее влияние аденогипофиза испытывают на себе Т- тироциты щитовидной железы, пучковая и сетчатая зоны коры надпочечников, эндокриноциты мужских и женских половых желез.
II. КЛАССИФИКАЦИЯ ЭНДОКРИННЫХ ЖЕЛЕЗ ПО ПРОИСХОЖДЕНИЮ (ИСТОЧНИКАМ РАЗВИТИЯ)
1. Эндокринные железы и клетки, развивающиеся из энтодермы.
а) бранхиогенная группа желез - эндокринные железы, происходящие
133
из эпителия глоточных карманов и близких к ним зачатков: Т-тироциты щитовидной железы, паращитовидные железы, тимус;
б) островки Лангерганса поджелудочной железы; в) одиночные эндокриноциты ДЭС пищеварительного тракта и возду-
хоносных путей (по другим представлениям, эндокриноциты ДЭС воздухоносных путей относятся к производным кожной эктодермы).
2.Эндокринные железы и клетки, развивающиеся из эктодермы и нейроэктодермы:
а) группа мозговых придатков - гипоталамус, гипофиз, эпифиз; б) мозговое вещество надпочечников, параганглии; в) кальцитониноциты щитовидной железы;
г) клетки Меркеля эпидермиса и эпителия волосяных фолликулов.
3.Эндокринные железы и клетки, развивающиеся из мезодермы и мезенхимы:
а) корковое вещество надпочечников; б) эндокриноциты половых желез; в) секреторные кардиомиоциты сердца; г) юкстагломерулярные клетки почки.
III. КЛАССИФИКАЦИЯ ПО УРОВНЮ СТРУКТУРНОЙ ОРГАНИЗАЦИИ
1)эндокринные органы (щитовидная и паращитовидные железы, надпочечник, гипофиз, эпифиз);
2)эндокринные части или ткани в составе органов, сочетающих эн-
докринные и неэндокринные функции (гипоталамус, островки Лангерганса поджелудочной железы, ретикулоэпителий и (возможно) тельца Гассаля тимуса, клетки Сертоли извитых семенных канальцев яичка и фолликулярный эпителий яичника);
3)клетки диффузной эндокринной системы (ДЭС).
ВЗАИМОСВЯЗЬ ЭНДОКРИННОЙ И НЕРВНОЙ СИСТЕМ
Как две регуляторные системы организма, эндокринная и нервная систем, тесно взаимодействуют при выполнении своих функций. У этих двух систем общий принцип регуляции функций клеток, тканей и органов, который заключается в синтезе и секреции регуляторных молекул - гормонов и нейромедиаторов. Разница между этими группами регуляторов не всегда резкая и заключается в том, что эндокринная система осуществляет гуморальную регуляцию в большинстве случаев по принципу эндокринии, тогда как нейроны выделяют свои нейромедиаторы в синаптические щели и действуют на контактирующие с ними другие клетки (ауто- и паракриния).
134
Наиболее тесная интеграция нервной и эндокринной систем осуществляется на уровне гипоталамуса. С одной стороны, гипоталамус является частью нервной системы, с другой - центральным органом эндокринной системы. В ядрах переднего и среднего гипоталамуса содержатся нейросекреторные нейроны, которые совмещают функции нервной и эндокринной клетки. Секретируемые ими нейрогормоны могут выступать в роли нейромедиаторов.
В последнее время установлена способность нейронов центральной нервной системы вырабатывать некоторые гормоны периферических эндокринных желез.
Выработка гормонов эндокринными железами тонко регулируется нервной системой. Каждая эндокринная железа богато иннервируется.
Примером тесной интеграции нервной и эндокринной систем является и мозговое вещество надпочечников, которое имеет единый источник развития с нервными ганглиями (ганглиозная пластинка) и продуцирует как гормон адреналин, так и нейромедиатор симпатической нервной системы
норадреналин.
Таким образом, тесное взаимодействие двух регуляторных систем - нервной и эндокринной - можно проследить на различных уровнях.
ГИПОТАЛАМУС
ФУНКЦИИ. Гипоталамус является центром регуляции вегетативных функций (надсегментарный центр симпатического и парасимпатическо-
го отделов ВНС) и высшим эндокринным центром. Он оказывает трансаденогипофизарное влияние (через стимуляцию выработки гипофизом тропных гормонов) на аденогипофиззависимые эндокринные железы и парааденогипофизарное влияние на аденогипофизнезависимые периферические эндокринные железы. Гипоталамус осуществляет контроль над всеми висцеральными функциями организма, объединяет нервные и эндокринные механизмы регуляции. Субстратом этого объединения являются нейросекреторные нейроциты, формирующие в гипоталамусе ядра. Вместе с гипофизом гипоталамус формирует структурно и функционально единую гипоталамо-
гипофизарную систему.
ИСТОЧНИКИ РАЗВИТИЯ. Источником развития гипоталамуса является нейроэктодерма - краниальный отдел нервной трубки. В результате миграционных процессов, совершаемых нейробластами II мозгового пузыря, дающего промежуточный мозг, ко 2-му месяцу эмбриогенеза формируются крупноклеточные, а к 3-4-му месяцу - большинство мелкоклеточных ядер. К 4-5-му месяцу дифференцируются нейроны крупноклеточных, а к 5-6-му месяцу - нейроны мелкоклеточных ядер.
135

СТРУКТУРНАЯ ОРГАНИЗАЦИЯ. Гипоталамус занимает базальную часть промежуточного мозга. Он находится под зрительным бугром (таламусом), окружая нижнюю часть 3-го желудочка головного мозга. Полость 3- го желудочка продолжается в воронку, направленную в сторону гипофиза. Стенка этой воронки называется гипофизарной ножкой. На дистальном конце она продолжается в заднюю долю гипофиза (нейрогипофиз). Кпере-
ди от гипофизарной ножки утолщение дна 3-го желудочка образует средин-
ное возвышение (медиальную эминенцию), содержащую первичную капиллярную сеть. В гипоталамусе выделяют передний, средний (медиобазальный) и задний отделы.
Основной частью гипоталамуса являются нервные и нейросекреторные клетки (нейроэндокриноциты). Они образуют более 30 (до 40) ядер (Рис. 16.2).
|
Рис. 16.2. Супраоптическое ядро гипо- |
||
|
таламуса. |
|
|
|
1 – |
крупные |
пептидхолинергические |
2 |
нейроны, формирующие ядро; |
||
|
2 – нервные волокна |
||
|
ПЕРЕДНИЙ |
ГИПОТАЛАМУС со- |
|
|
держит наиболее крупные парные |
||
|
супраоптические и паравентрику- |
||
|
лярные ядра. Кроме них, здесь на- |
||
|
ходится ряд других ядер серого ве- |
||
|
щества гипоталамуса. Супраоптиче- |
||
|
ские ядра образованы в основном |
||
1 |
крупными пептидхолинергически- |
||
|
ми нейронами (нейроэндокрино- |
||
|
цитами). В их цитоплазме выявля- |
||
2 |
ются |
нейросекреторные гранулы и |
|
|
хроматофильная субстанция Ниссля. |
||
|
При |
электронномикроскопическом |
|
исследовании в пептидхолинергических нейронах выявляются сильно развитый белоксинтезирующий и секреторный аппараты, нейросекреторные гранулы и синаптические пузырьки, что свидетельствует о параллельном образовании нейрогормонов и нейромедиаторов. На перикарионе нейросекреторных клеток образуется большое количество интернейрональных синапсов, свидетельствующих о включении данных клеток в рефлекторные дуги. Аксоны пептидхолинергических нейроцитов через гипофизарную ножку на-
136

правляются в заднюю долю гипофиза и образуют там синапсы на кровеносных сосудах (нейрососудистые синапсы, Рис. 16.3). Нейроны супраоптиче-
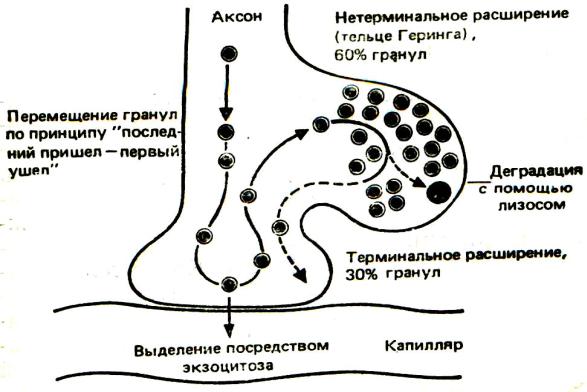
ских ядер секретируют в основном антидиуретический гормон (АДГ), или вазопрессин, в меньшем количестве - окситоцин. Эти гормоны с помощью аксотока транспортируются по аксонам в заднюю долю гипофиза и накапливаются в расширениях аксонов, которые лежат выше нейрососудистого си-
напса и называются претерминальными расширениями аксона (накопительными тельцами Херринга). При необходимо-
Рис. 16.3. Гипоталамогипофизарная система и действие тропных гормонов на органымишени (по Б.В. Алешину).
1 – зрительный перекрест; 2 - медиальное возвышение с первичной капиллярной сетью; 3 – полость III желудочка, проекция некоторых ядер на стенку этого желудочка; 4 –супраоптическое ядро; 5 – переднее гипоталамическое ядро (преоптическая зона гипоталамуса; 6 – паравентрикулярное ядро; 7
– аркуатно-медиобазальный комплекс медиобазального гипоталамуса; 8 – нейросекреторные пептидадренергические нейроны медиобазального гипоталамуса, секретирующие рилизинг-факторы в первичную капиллярную сеть; 9 – адренергические нейроны медиобазального гипоталамуса, дающие начало нисходящим эфферентным путям (парагипофизарная передача гипоталамических импульсов регулируемым эффекторам; 10 – таламус; 11 – воронка III желудочка и гипофизарная ножка; 12 – задняя доля гипофиза; 13 – накопительное тельце Херринга (окончания аксонов нейросекреторных нейронов переднего гипоталамуса – супраоптического и
паравентрикулярного ядер - на капиллярах задней доли гипофиза); 14 – средняя доля гипофиза; 15 – гипофизарная щель; 16 – передняя доля гипофиза со вторичной капиллярной сетью; 17 – портальная (воротная) вена; 18 – туберальная часть аденогипофиза. Аденогипофизарные гормоны: СТГ – соматотропный гормон; ЛГ – лютеинизирующий гормон, лютропин; ФСГ – фолликулостимулирующий гормон,
137
фоллитропин; ТТГ – тиротропный гормон, тиротропин; ЛТГ – лактотропный гормон, лактотропин, ммаммотропин Гормоны, содержащиеся в задней доле гипофиза: Окс – окситоцин; АДГ – антидиуретический гормон. Э – эстрогены; ПГ – прогестерон
сти гормоны из накопительного тельца Херринга поступают к нейрогемальным синапсам, где имеются терминальные расширения аксона, а затем в кровь (Рис. 16.4). В тельцах Херринга депонируется около 60%, в терминальных расширениях аксона – 30% всех гранул. Органамимишенями АДГ являются почки и артериолы. В почках он усиливает реабсорбцию воды в кровь (это происходит в канальцах нефрона и собирательных трубочках, в которых вазопрессин действует на аквапорины - водные каналы, увеличивая их проницаемость для воды) и тем самым уменьшает объем мочи, вызывая задержку жидкости в организме, увеличение объема циркулирующей жидкости, повышение артериального давления и снижение диуреза. Этим эффектом обусловлено первое название гормона - АДГ.
Рис. 16.4. Перемещение и накопление нейросекреторных гранул в аксонных расширениях и связанных с капиллярами терминалях крупноклеточных нейронов в задней доле гипофиза. Относительное распределение гранул между аксонами, субтерминальными расширениями (тельцами Херринга) и терминальными расширениями аксона (по Дж. Моррис)
В артериолах гормон вызывает сокращение гладких миоцитов мышечной оболочки и повышение АД, отсюда его второе название (вазопрессин). При генетически обусловленных нарушениях выработки АДГ в гипоталамусе или вследствие дефекта в почках рецепторов к нему (аквапоринов) реаб-
138