

1 Переваривание нуклеиновых кислот и нуклеотидов в желудочно- кишечном тракте.
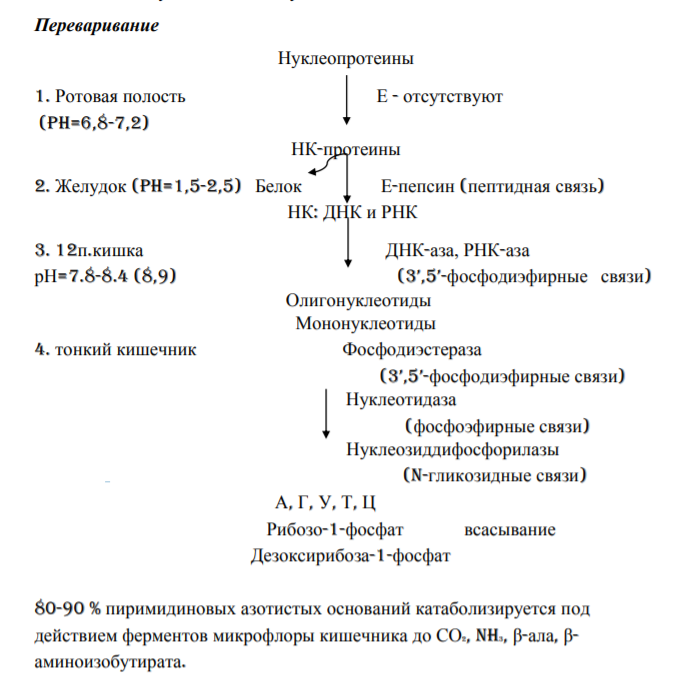
2 Источники атомов углерода и азота пуринового кольца.
Печень - основное место образования пуриновых нуклеотидов, откуда они могут поступать в ткани, не способные к их синтезу: эритроциты, ПЯЛ и частично мозг.
Источники атомов углерода и азота в пуриновом кольце:
1-из аспарагиновой кислоты
2- из формил - ТГФК
3,9- из глутамина
4,5,7 - из глицина
6- из СО2
8- из метенил – ТГФК
3 Схема биосинтеза АМФ и ГМФ de novo.
«de novo» основной путь 80-90%; локализация – печень
1. Субстраты для синтеза пурина:
СO2 + гли + глн + асн + N10-формил Н 4-фолат + N10-метенил Н4-фолат
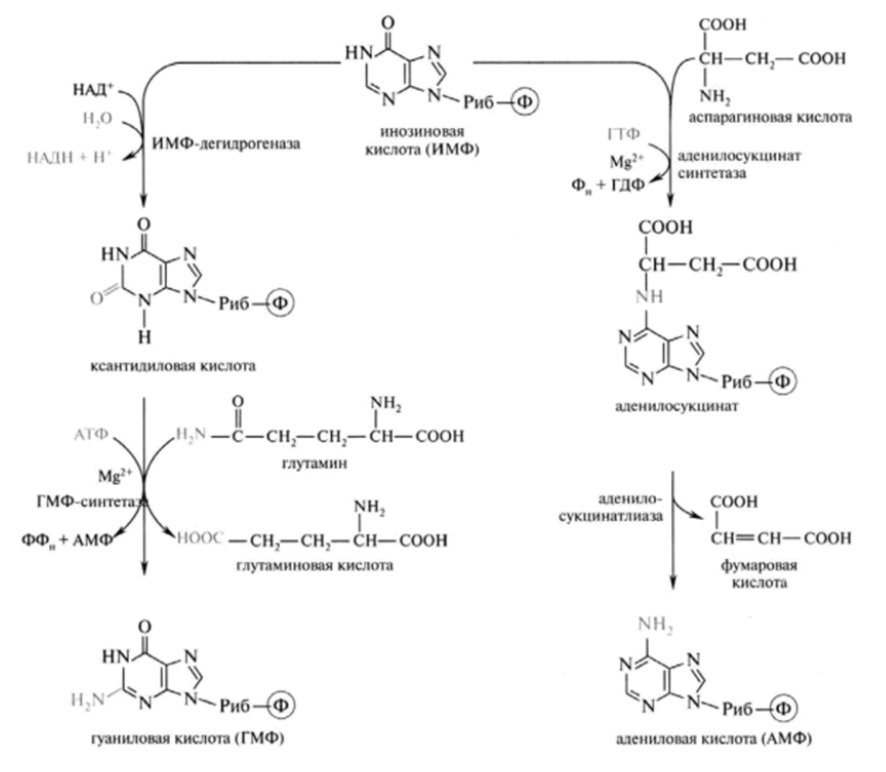
4. Резервный путь биосинтеза АМФ, ГМФ Схема синтеза ИМФ АМФ, ИМФ, ГМФ
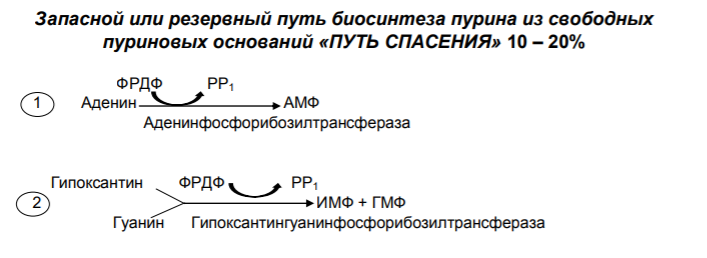
5. Нарушения обмена пуриновых нуклеотидов в организме человека (подагра) .
Вследствие низкой (малой) растворимости мочевой кислоты, в моче присутствуют ее соли – ураты (которые более растворимы). Большинство камней в мочевыводящей системе – кристаллы мочевой кислоты.
Повышение концентрации мочевой кислоты в плазме крови приводят к гиперурикемии, следствием которой является подагра – заболевание, при котором кристаллы мочевой кислоты и уратов откладываются в суставных хрящах синовиальной оболочке, подкожной клетчатке с образованием подагрических узлов, или тофусов. Признаки подагры – острое воспаление суставов - острый подагрический артрит.
Поскольку лейкоциты фагоцитируют кристаллы уратов, то причиной воспаления является разрушение лизосомальных мембран лейкоцитов кристаллами мочевой кислоты. Освободившиеся лизосомальные ферменты выходят в цитозоль и разрушают клетки, а продукты клеточного катаболизма вызывают воспаление.
6. Схема биосинтеза пиримидиновых нуклеотидов в тканях.
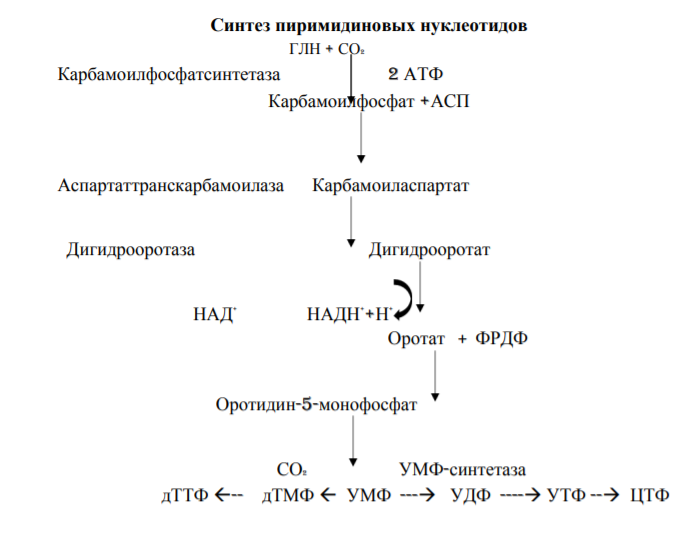

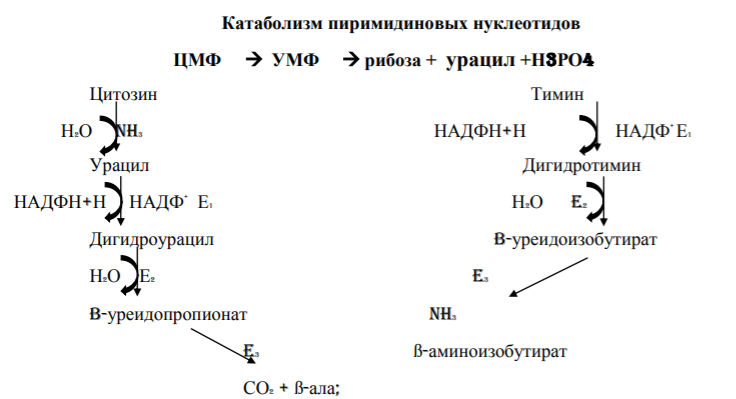
7. Распад пиримидиновых нуклеотидов в тканях.
1.Матричные биосинтезы в организме: репликация, репарация (биосинтез дНк); транскрипция (биосинтез м-рнк, р-рнк, т-рНк). Трансляция (биосинтез белка). Общая характеристика.
Репликация – синтез ДНК, где каждая из двух цепей служит матрицей для образования новой цепи. Источниками энергии являются дезоксирибонуклеозидтрифосфаты (дНТФ: дАТФ, дГТФ, дЦТФ, дТТФ).
Транскрипция – синтез РНК на ДНК-матрице, образуются первичные транскрипты – мРНК, тРНК, рРНК, комплементарны матричной цепи ДНК, имеющей направление от 3՛ к 5՛-концу. Субстратами и источниками энергии для синтеза РНК являются рибонуклеозидтрифосфаты (НТФ: УТФ, ГТФ, ЦТФ, АТФ).
Трансляция – непосредственно синтез белка. Информация записана на четырёх буквенном языке нуклеиновых кислот (ТАГЦ), переводится на язык белков, состоящий из 20 букв. Синтез белка отличается от других матричных биосинтезов (репликации, транскрипции) тем, что между мРНК и продуктом-белком нет комплиментарного соответствия.
Репарация ошибок и повреждений ДНК– восстановление ДНК, основанное на том, что ДНК - двухцепочечная молекула. Если нуклеотидная последовательность одной из двух цепей оказывается повреждённой (изменённой), информацию можно восстановить, так как вторая (комплементарная) цепь сохранена
2. Pепликация днк, характеристика процесса, механизм, субстраты, этапы, ферменты, биологическое значение
Репликация– это синтез, удвоение цепей ДНК, включает три основных этапа:
I. Формирование репликативной вилки.
II. Синтез новых цепей ДНК.
III. Исключение праймеров и завершение формирования отстающей цепи ДНК.
I. Формирование репликативной вилки.Основные ферменты:
1. ДНК – топоизомеразы разрывают 3՛-5՛-фосфодиэфирную связь одной из цепей ДНК и присоединяется к 5՛ в точке разрыва, образуется репликативная вилка.
2. ДНК-хеликазы – присоединяются в области разрыва двух цепей, также участвуя в образовании репликативной вилки, т.о. расплетают двойную спираль ДНК.
3. SSB (singstrandbinding) – белки, связываются с одноцеполчечными участками ДНК, предотвращая повторное комплементарное скручивание.
II. Синтез новых цепей ДНК
I этап – дочерние нити образуются на обеих нитях материнской ДНК. Основные ферменты: ДНК-полимеразы, они синтезируют полинуклеотидные цепи из нуклеотидтрифосфатов (дАТФ, дГТФ, дТТФ, дЦТФ) в направлении от 5՛к 3՛-концу антипараллельно матрице, имеющей направление от 3՛ к 5՛- концу.
III этап синтеза ДНК – исключение праймеров и завершение формирования отстающей цепи ДНК.
1. В отстающей нити праймер удаляется эндонуклеазой.
2. ДНК-полимераза β заполняет образованную «брешь», присоединяя дезоксирибонуклеотиды.
3. Присоединение дезоксирибонуклеотида путём связывания 3՛-ОН одного фрагмента с 5՛-фосфатом предыдущего фрагмента; катализирует ДНК-лигаза. Это процесс, когда из множества фрагментов Оказаки образуется нить ДНК.
4. Результатом является образование дочерних цепей комплементарных и антипараллельных нитям материнской ДНК.
5. Молекулы ДНК человека имеют очень большие размеры, поэтому инициация синтеза ДНК происходит в точках инициации репликации ДНК – ориджинах репликации. Синтез ДНК начинается в области ориджина, идет в противоположных направлениях, в каждом ориджине образуются две репликативные вилки, которые перемещаются в противоположных направлениях, пока не встретятся.
6. Единица репликации у эукариот называется репликоном. Репликон– это участок ДНК между соединенными ориджинами.