

23. Строение и значение пероксисом

Пероксисомы (микротельца) по строению сходны с лизосомами. Они состоят из матирикса и нуклеотида. Матрикс пероксисом содержит до 15 ферментов. Наиболее важные из них пероксидаза и каталаза, оксидаза D-аминокислот и уратроксидаза. Нуклеотид пероксисомы соответствует области конденсации ферментов. Пероксисомы образуются в ЭПС, отпочковываясь от агранулярной ЭПС, их ферменты частично синтезируются в гранулярной ЭПС, частично в гиалоплазме. Мембрана пероксисом непроницаема для ионов и низкомолекулярных субстратов.
+Пероксисомы – главный центр образования кислорода клетки. В результате окисления аминокислот, углеводов образуется Н2О2, которая благодаря каталазе распадается на воду и О2. Крупные пероксисомы печени и почек играют важную роль в обезвреживании ряда веществ. Помимо этого они участвуют в катаболизме (в обмене аминокислот, оксалата и полиаминов).
В настоящее время открыт класс наследственных болезней – пероксисомные болезни, развитие которых обусловлекнно дефектом пероксисом. При этих болезнях поражаются органы, развиваются нарушения нервной системы, вызывающих смерть больных в детском возрасте.
24. Строение и значение рибосом.
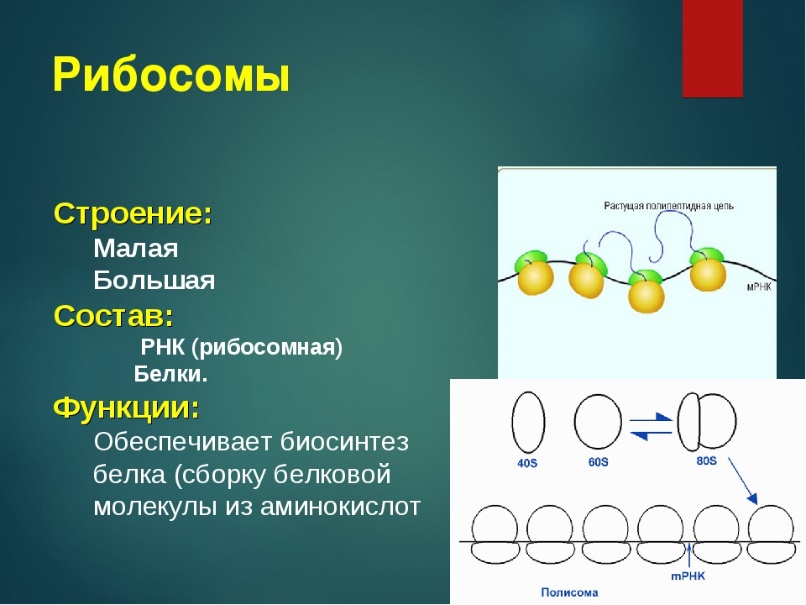

Рибосомы, внутриклеточные частицы, осуществляющие биосинтез белка В процессе функционирования (т. е. синтеза белка) Рибосомы осуществляет несколько функций:
1) специфическое связывание и удержание компонентов белоксинтезирующей системы [информационная, или матричная, РНК (иРНК) : аминоацил-тРНК; пептидил-тРНК; гуанозинтрифосфат (ГТФ) ; белковые факторы трансляции EF - Т и EF - G]:
2) каталитические функции (образование пептидной связи, гидролиз ГТФ):
3) функции механического перемещения субстратов (иРНК, тРНК) , или транслокации. Функции связывания (удержания) компонентов и катализа распределены между двумя рибосомными субчастицами. Малая рибосомная субчастица содержит участки для связывания иРНК и аминоацил-тРНК и, по-видимому, не несёт каталитических функций. Большая субчастица содержит каталитический участок для синтеза пептидной связи, а также центр, участвующий в гидролизе ГТФ: кроме того, в процессе биосинтеза белка она удерживает на себе растущую цепь белка в виде пептидил-тРНК. Каждая из субъединиц может проявить связанные с ней функции отдельно, без связи с другой субчастицей. Однако ни одна из субчастиц в отдельности не обладает функцией транслокации, осуществляемой только полной Рибосомы
25. Цитоскелет клетки, его структурные элементы и их производные.


Цитоскелет - это совокупность нитевидных белковых структур находящихся в цитоплазме живой клетки и образующих клеточный скелет или каркас.
Цитоскелет представляет собой сложную динамичную систему микротрубочек, микрофиламентов, промежуточных филаментов и микротрабекул. Указанные компоненты цитоскелета являются немембранными органеллами; каждый из них образует в клетке трехмерную сеть с характерным распределением, которая взаимодействует с сетями из других компонентов. Они входят также в состав ряда других более сложно организованных органелл (ресничек, жгутиков, микроворсинок, клеточного центра) и клеточных соединений (десмосом, полудесмосом, опоясывающих десмосом).
Основные функции цитоскелета:
1.поддержание и изменение формы клетки;
2.распределение и перемещение компонентов клетки;
3.транспорт веществ в клетку и из нее;
4.обеспечение подвижности клетки;
5.участие в межклеточных соединениях.
Микротрубочки - наиболее крупные компоненты цитоскелета. Они представляют собой полые цилиндрические образования, имеющие форму трубочек, длиной до нескольких микрометров (в жгутиках более 50 им) диаметром около 24-25 нм, с толщиной стенки 5 нм и диаметром просвета 14-15 нм (рис. 3-14).
Стенка микротрубочки состоит из спиралевидно уложенных нитей - протофиламентов толщиной 5 нм (которым на поперечном разрезе соответствуют 13 субъединиц), образованных димерами из белковых молекул α- и β-тубулина.
Функции микротрубочек:
1.поддержание формы и полярности клетки, распределения ее компонентов,
2.обеспечение внутриклеточного транспорта.
3.обеспечение движения ресничек, хромосом в митозе (формируют ахроматиновое веретено, необходимое для клеточного деления),
4.образование основы других органелл (центрнолей, ресничек).
Расположение микротрубочек. Микротрубочки располагаются в цитоплазме в составе нескольких систем:
а) в виде отдельных элементов, разбросанных по всей цитоплазме и формирующих сети;
б) в пучках, где они связаны тонкими поперечными мостиками (в отростках нейронов, в составе митотического веретена, манжетки сперматиды, периферического "кольца" тромбоцитов);
в) частично сливаясь друг с другом с формированием пар, или дyблетов (в аксонеме ресничек и жгутиков), и триплетов (в базальном тельце и центриоли).
Образование и разрушение микротрубочек. Микротрубочки представляют собой лабильную систему, в которой имеется равновесие между их постоянной сборкой и диссоциацией. У большинства микротрубочек один конец (обозначаемый как "-") закреплен, а другой ("+") свободен и участвует в их удлинении или деполимеризации. Структурами, обеспечивающими образование микротрубочек, служат особые мелкие сферические тельца - сателлиты (от англ. satellite - спутник), отчего последние называют центрами организации микротрубочек (ЦОМТ). Сателлиты содержатся в базальных тельцах ресничек и клеточном центре (см. рис. 3-15 и 3-16). После полного разрушения микротрубочек в цитоплазме они отрастают от клеточного центра со скоростью около 1 мкм/мин., а их сеть вновь восстанавливается менее, чем за полтора часа. К ЦОМТ относят также и центромеры хромосом.
Связь микротрубочек с другими структурами клетки и между собой
осуществляется посредством ряда белков, выполняющих различные функции.
(1) Микротрубочки с помощью вспомогательных белков прикреплены к другим клеточным компонентам. (2) По своей длине микротрубочки образуют многочисленные боковые выросты (которые состоят из белков, ассоциированных с микротрубочками) длиной до нескольких десятков нанометров. Благодаря тому, что такие белки последовательно и обратимо связываются с органеллами, транспортными пузырьками, секреторными гранулами и другими образованиями, микротрубочки (которые сами не обладают сократимостью) обеспечивают перемещение указанных структур по цитоплазме. (3) Некоторые белки, ассоциированные с микротрубочками, стабилизируют их структуру, а связываясь с их свободными краями, препятствуют деполимеризации.
Клеточный центр образован двумя полыми цилиндрическими структурами длиной 0.3-0.5 мкм и диаметром 0.15-0.2 мкм - центриолями, которые располагайся вблизи друг друга во взаимно перпендикулярных плоскостях (рис. 3-15). Каждая центриоль состоит из 9 триплетов частично слившихся микротрубочек (А, В и С), связанных поперечными белковыми мостиками ("ручками"). В центральной часта центриоли микротрубочки отсутствуют (по некоторым данным, здесь имеется особая центральная нить), что описывается общей формулой (9*3) + 0. Каждый триплет центриоли связан со сферическими тельцами диаметром 75 нм - сателлитами-, расходящиеся от них микротрубочки образуют центросферу.
В неделящейся клетке выявляется одна пара центриолей (диплосома), которая обычно располагается вблизи ядра. Перед делением в S-периоде интерфазы происходит дупликация центриолей пары, причем под прямым углом к каждой зрелой (материнской) центриоли формируется новая (дочерняя), незрелая процентриоль, в которой вначале имеются лишь 9 единичных микротрубочек, позднее превращающихся в триплеты. Пары ценгриолей далее расходятся к полюсам клетки, а во время митоза они служат центрами образования микротрубочек ахроматинового веретена деления.
Микрофиламенты - тонкие белковые нити диаметром 5-7 нм, лежащие в цитоплазме поодиночке, в виде сетей ими пучками. В скелетной мышце тонкие микрофиламенты образуют упорядоченные пучки, взаимодействуя с более толстыми миознновыми филаментами.
Кортикальная (терминальная) сеть - зона сгущения микрофиламентов под плазмолеммой, характерная для большинства клеток. В этой сети микрофиламенты переплетены между собой и "сшиты" друг с другом с помощью особых белков, самым распространенным из которых является филамин. Кортикальная сеть препятствует резкой и внезапной деформации клетки при механических воздействиях и обеспечивает плавные изменения ее формы путем перестройки, которая облегчается актин-растворяющими (преобразующими) ферментами.
Прикрепление микрофиламентов к плазмолемме осуществляется благодаря их связи с её интегральными ("якорными") белками (интегринами) - непосредственно или через ряд промежуточных белков - талин, винкулин и α- актинин (см. рис. 10-9). Помимо этого, актиновые микрофиламенты прикрепляются к трансмембранным белкам в особых участках плазмолеммы. называемых адгезионными соединениями, или фокальными контактами, которые связывают клетки друг с другом или клетки с компонентами межклеточного вещества.
Актин - основной белок микрофиламентов - встречается в моно-мерной форме (G-, или глобулярный актин), которая способна в присутствии цАМФ и Са2+ полимеризоваться в длинные цепи (F-, или фибриллярный актин). Обычно молекула актина имеет вид двух спирально скрученных нитей (см. рис. 10-9 и
В микрофиламентах актин взаимодействует с рядом актин-связывающих белков (до нескольких десятков видов), выполняющих различные функции. Некоторые из них регулируют степень полимеризации актина, другие (например, филамин в кортикальной сети или фимбрин и виллин в микроворсинке) способствуют связыванию отдельных микрофиламентов в системы. В немышечных клетках на актин приходится примерно 5-10% содержания белка, лишь около половины его организовано в филаменты. Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Функции микрофиламентов:
(1)обеспечение сократимости мышечных клеток (при взаимодействии с миозином);
(2)обеспечение функций, связанных с кортикальным слоем цито-плазмы и плазмолеммой (экзо- и эндоцитоз, образование псевдоподий и миграция клетки);
(3)перемещение внутри цитоплазмы органелл, транспортных пузырьков и других структур благодаря взаимодействию с некоторыми белками (минимиозином), связанными с поверхностью этих структур;
(4)обеспечение определенной жесткости клетки за счет наличия кортикальной сети, которая препятствует действию деформаций, но сама, перестраиваясь, способствует изменениям клеточной формы;
(5)формирование сократимой перетяжки при цитотомии, завершающей клеточное деление;
(6)образование основы ("каркаса") некоторых органелл (микро-ворсинок, стереоцилий).
(7)участие в организации структуры межклеточных соединений
(опоясывающих десмосом).
Промежуточные филаменты - прочные и устойчивые в химическом отношении белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микро-филаментов). Они встречаются в клетках разных тканей (см. ниже) и располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, входят в состав десмосом и полудесмосом эпителиальных клеток (в плазмолемме которых они закреплены посредством трансмембранных белков), лежат по всей длине отростков нейронов. Промежуточные филаменты образованы нитевидными белковыми молекулами, сплетенными друг с другом наподобие каната.
Функции промежуточных филаментов изучены недостаточно;
установлено, однако, что они не влияют ни на движение, ни на деление клетки.
Ких основным функциям относятся:
(1)структурная - поддерживающая и опорная, обеспечение распределения органелл по определенным участкам цитоплазмы;
(2)обеспечение равномерного распределения сил деформации между клетками ткани, что препятствует повреждению отдельных клеток (благодаря связи промежуточных филаментов с трансмембранньгми белками десмосом и полудесмосом);
(3)участие в образовании рогового вещества в эпителии кожи
(4)поддержание формы отростков нервных клеток и фиксация трансмембраниых белков (в частности, ионных каналов);
(5)удержание миофибрилл в мышечной ткани и прикрепление их к плазмолемме. что обеспечивает их сократительную функцию.