

56. Амфиболические функции цикла трикарбоновых кислот.
57. Анаболические функции цикла лимонной кислоты. Реакции, пополняющие цитратный цикл.
Амфиболические ф-ции ЦТК – катаболическая ф-ция, обеспечение распада ацетата и следоват. спос. катаболизму углеводов, липидов, ам-т, с др. стороны субстраты цикла исп. в анаболич. целях - для синтеза глюкозы, дикарбоновых к-т. порфиринов.
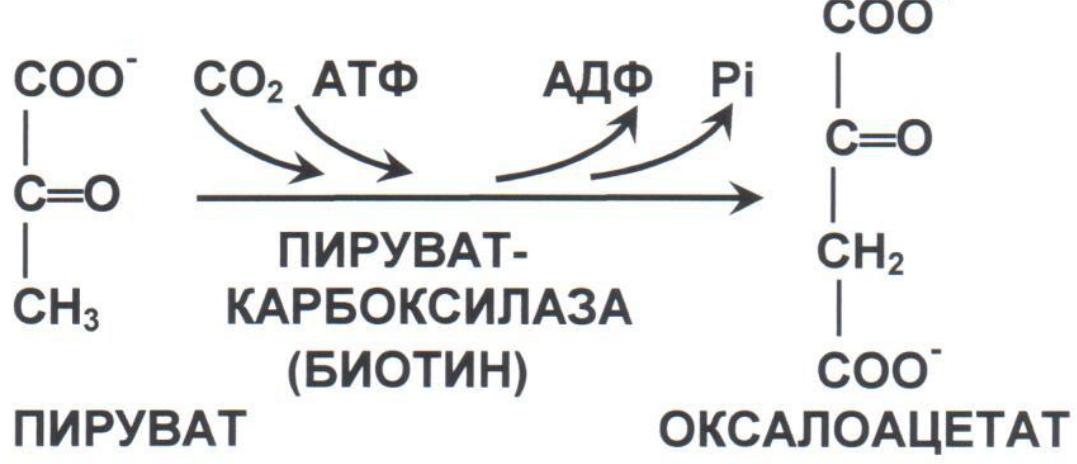
Анаплероз-процесс
восполнеия резерва промежуточных
продуктов цикла.Анаплеротические р-ции
– ферментативные реакции образования
пирувата и промежут.продуктов цикла.
Основная реакция — превращение
пировиноградной кислоты в ацетоацетат.
54. Представление о процессах окислительного фосфорилирования. Надн-дегидрогеназа как компонент дыхательной цепи.
57. Дыхательная цепь митохондрий, ее строение и основные принципы функционирования. Теории сопряжения окисления и фосфорилирования
Окислительное фосфолирование – процесс образования АТФ и воды на внут.мембране митохондрий, заключающийся в окислении восстановленных эквивалентов (НАДН и ФАДН2) дыхательными ферментами.
На внут.мембране содержится мультиферментных комплекс-дыхательные ферменты, а последовательность их расположения в мембране назыв.дыхат.цепью.
Дыхат. цепь состоит из: НАД-зависимой дегидрогеназы; ФАД-зависимой дегидрогеназы; Убихинона (КоQ); Цитохрмов b, c, a+a3. Соединение может отдавать электроны только соед-ю с более высоким ок-восст потенциалом. В дых. цепи каждое последующее звено имеет более высокий потенциал, чем предыдущее.
НАДН дегидрогеназа окисляет НАД-Н, отбирая у него два электрона и перенося их на растворимый в липидах убихинон, который внутри мембраны диффундирует к комплексу III. Вместе с этим, комплекс I перекачивает 4 протона из матрикса в межмембранное пространство митохондрии.
58-59. Окислительное декарбоксилирование ПВК. Химизм процесса и его биологическое значение
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных в «пируватдегидрогеназный комплекс»:3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 коферментов (тиамидфасфат(кофермент В6), липоевая кислота, КоА(кофермент пантотеновой к-ты), ФАД(кофермент В2) и НАД(В3).
На
I стадии этого процесса пируват теряет
свою карбоксильную
группу
в результате взаимодействия с
тиаминпирофосфатом (ТПФ) в составе АЦ
фермента
пируватдегидрогеназы.
 60.
Окислительное фосфорилирование,
коэффициент Р/О. Строение митохондрий
и структурная организация дыхательной
цепи. Трансмембранный электрохимический
потенциал как промежуточная форма
энергии при окислительном фосфорилировании.
60.
Окислительное фосфорилирование,
коэффициент Р/О. Строение митохондрий
и структурная организация дыхательной
цепи. Трансмембранный электрохимический
потенциал как промежуточная форма
энергии при окислительном фосфорилировании.
Коэфф. фосфолирования Р/О - соотношение колич. израсход. на синт. АТФ Н3РО4 и поглощенного О2. Выраж. эффективность функционирования цепи транспорта эл., чем выше этот коэфф. , тем больше синт-ся АТФ в расчете на каждую пару синт-х электронов. В случае полной дых. цепи равен 3.
Трансмембр. электрохимич. потенциал. - градиент конц. ионов водорода и эл. зарядов по обес строноны мембр. митох..Избыт. величина электрохим потенциала необх для обесп. транспорта из цитоплазмы в митохондрию ПВК, неорг. фосфата, АДФ, а из митох в цитопл. - АТФ.
61-63. Токсичность кислорода: образование активных форм кислорода (супероксид анион, перекись водорода, гидроксильный радикал). Повреждение мембран в результате перекисного окисления липидов. Защита от токсического действия кислорода: неферментативные и ферментативные антиоксиданты.
О2,
необх. организму для функ-ния ЦПМ и
многих других реакций, является
одновременно и токсическим веществом,
если из него образуются так называемые
активные формы: ОН• - гидроксильный
радикал;![]() - супероксидный анион;Н2О2 - пероксид
водорода. Наиболее активен гидроксильный
радикал, взаимодействующий с большинством
органических молекул. Он отнимает от
них электрон и инициирует таким образом
цепные реакции окисления. Эти
свободнорадикальные реакции окисления
могут выполнять полезные функции,
например, когда клетки белой крови с
участием активных форм кислорода
разрушают фагоцитированные клетки
бактерий. Но в остальных клетках
свободнорадикальное окисление приводит
к разрушению органических молекул, в
первую очередь липидов, и, соответственно,
мембранных структур клеток, что часто
заканчивается их гибелью. Поэтому в
организме функционирует эффективная
система ингибирования перекисного
окисления липидов (ПОЛ).
- супероксидный анион;Н2О2 - пероксид
водорода. Наиболее активен гидроксильный
радикал, взаимодействующий с большинством
органических молекул. Он отнимает от
них электрон и инициирует таким образом
цепные реакции окисления. Эти
свободнорадикальные реакции окисления
могут выполнять полезные функции,
например, когда клетки белой крови с
участием активных форм кислорода
разрушают фагоцитированные клетки
бактерий. Но в остальных клетках
свободнорадикальное окисление приводит
к разрушению органических молекул, в
первую очередь липидов, и, соответственно,
мембранных структур клеток, что часто
заканчивается их гибелью. Поэтому в
организме функционирует эффективная
система ингибирования перекисного
окисления липидов (ПОЛ).
Среда с сод-м кислорода явл. агрессивной по Антиоксиданты - ингибиторы окисления прир. или синтетич. в-ва. спос. тормозить окисление. Мех-м действия сост. в обрыве реакционных цепей, молек. антиокс. взаимод с акт. радик. с обр. малоакт. радик., уменьш скорость окисления