ПРАКТИЧЕСКОЕ ЗАНЯТИЕ №2
ТЕМА: РЕГУЛЯЦИЯ ОНТОГЕНЕЗА. МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЕ МЕХАНИЗМЫ ДИФФЕРЕНЦИРОВКИ. КРИТИЧЕСКИЕ ПЕРИОДЫ. РОЛЬ СРЕДЫ В РАЗВИТИИ ОРГАНИЗМА.
Вопросы для самоподготовки:
1. Основные концепции в биологии развития: преформизм и эпигенез.
2. Характеристика клеточных механизмов развития:
Деление
Миграция
Сортировка
запрограммированная гибель
адгезия
дифференцировка (лабильная и стабильная)
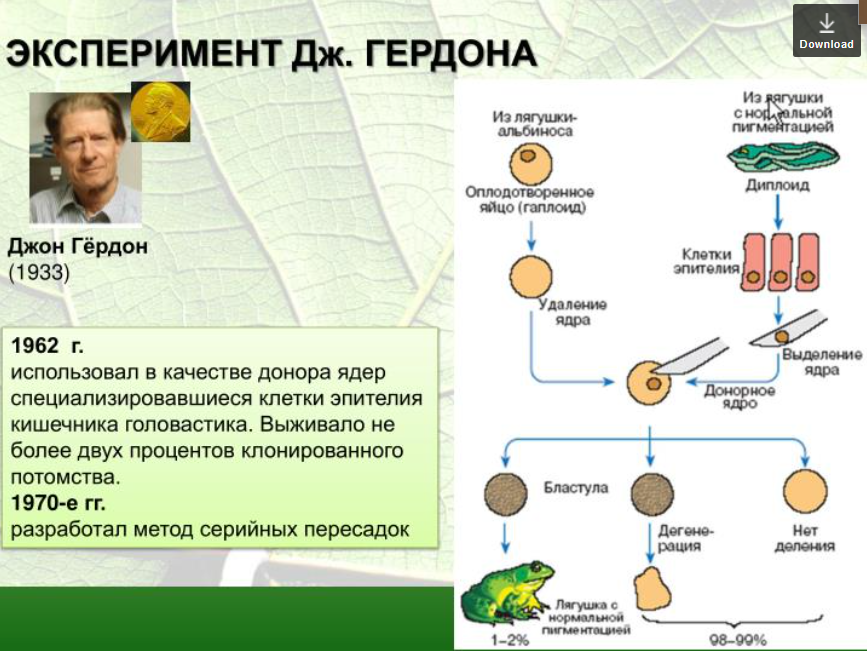
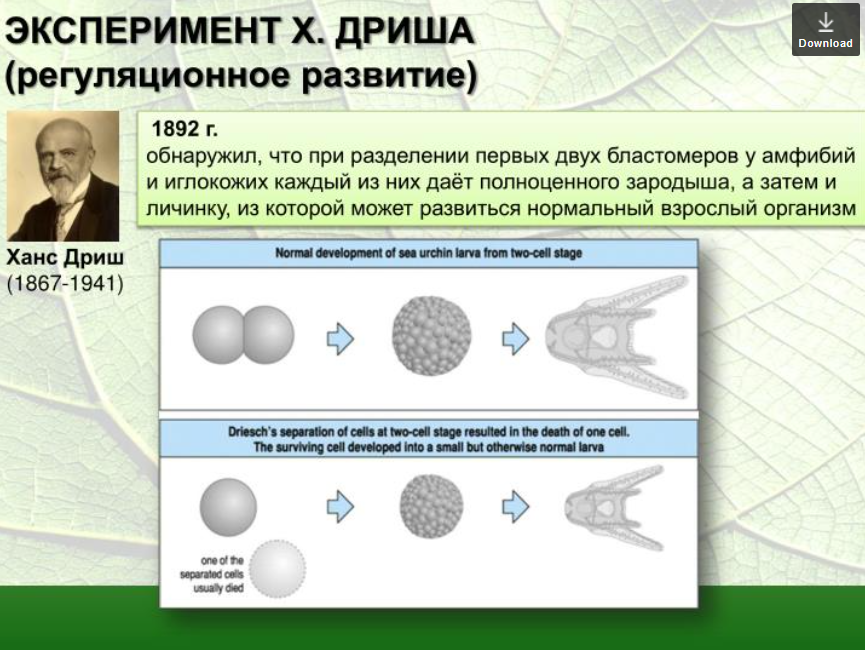
3. Методы экспериментальной эмбриологии. Опыты В. Ру и Д. Гердона, Г. Шпемана, Г. Дриша по изучению закономерностей эмбриогенеза.
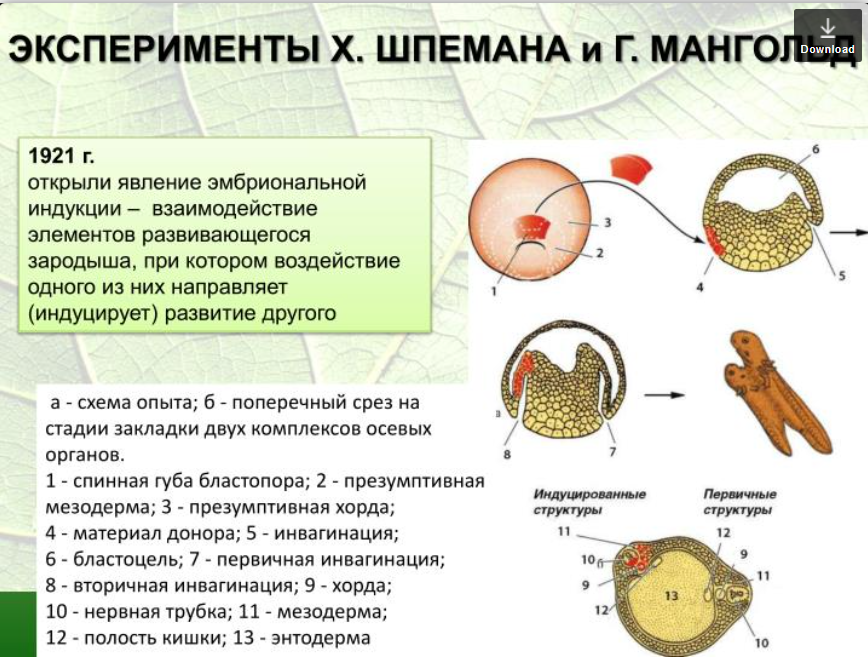
4. Роль эмбриональной индукции в морфообразовательном процессе. Опыты Г.Шпемана.
5. Генетический контроль развития:
гомеозисные и дизруптивные мутации
мутации генов с материнским эффектом
мутации генов Т-локуса на примере мыши
6. Гормональная регуляция онтогенеза на примере плацентарных млекопитающих.
7. Целостность онтогенеза. Понятие детерминации в развитии.
8. Роль эмбриональной регуляции в сохранении нормального хода развития.
9. Понятие морфогенеза. Концепции регуляции морфогенеза:
физиологических градиентов
позиционной информации
морфогенетических полей
10. Понятие критических периодов в онтогенезе человека.
11. Определение и классификация врождённых пороков развития.
1. Основные концепции в биологии развития: преформизм и эпигенез.
Преморфизм – в этой теории онтогенез рассматривали лишь как рост расположенных в определенном пространственном порядке предсуществующих структур и частей будущего организма. В этих рамках каких-либо новообразований или преобразований структур в индивидуальном развитии не происходит. Логическое завершение идеи преформизма заключается в допущении абсурдной мысли о ≪заготовленности≫ в зиготе и даже в половых клетках прародителей структур организмов всех последующих поколений, как бы вложенных последовательно наподобие деревянных матрешек.
Эпигенез – альтернатива периморфизму. сформулирована в середине XVIII в. Ф. К. Вольфом, впервые обнаружившим новообразование нервной трубки и кишечника в ходе эмбрионального развития. Индивидуальное развитие стали связывать целиком с качественными изменениями, полагая, что структуры и части организма возникают как новообразования из бесструктурной яйцеклетки.
В XIX в. К. Бэр впервые описал яйцо млекопитающих и человека, а также зародышевые листки и обнаружил сходство плана строения зародышей различных классов позвоночных — рыб, амфибий, рептилий, птиц, млекопитающих. Он же обратил внимание на преемственность в этапах развития — от более простого к более сложному. Бэр рассматривал онтогенез не как предобразование, не как новообразование структур, а как их преобразование, что вполне согласуется с современными представлениями.
2. Характеристика клеточных механизмов развития:
Элементарные клеточные механизмы онтогенеза
В онтогенезе особи происходят сложнейшие преобразования: осуществляется дифференциация частей развивающегося организма (морфогенез), формирование его внешней и внутренней структуры (морфогенез), рост. В основе этих преобразований лежат клеточные и системные механизмы развития. К клеточным механизмам относят размножение, перемещения, избирательную сортировку, дифференцировку, программированную гибель клеток.
А) Деление
Деление клеток (размножение, пролиферация) играет важную роль в процессах онтогенеза. Во-первых, благодаря делению из зиготы, которая соответствует одноклеточной стадии развития, возникает многоклеточный организм. Во-вторых, пролиферация клеток, происходящая после стадии дробления, обеспечивает рост организма. В-третьих, избирательному размножению клеток принадлежит заметная роль в обеспечении морфогенетических процессов. В-четвертых, в постнатальном периоде индивидуального развития благодаря клеточному делению осуществляется обновление многих тканей в процессе жизнедеятельности организма (физиологическая или гомеостатическая регенерация), а также заживление ран, восстановление утраченных органов (репаративная регенерация)
Б) Миграция
В процессе развития особи происходят неоднократные миграции отдельных клеток, их групп, клеточных пластов. Особое значение миграция клеток приобретает на стадии гаструляции, приводя к формированию зародышевых листков. В ходе органогенеза этот механизм важен, например, при формировании крупных пищеварительных желез, производных нервного гребня. Не менее значима его роль и в постэмбриональном развитии. Амебоидное движение макрофагов обеспечивает реализацию реакций иммунитета, перемещения сперматозоидов (жгутиковое движение) необходимы для осуществления оплодотворения, миграции клеток эпидермиса приводят к закрытию раневой поверхности при повреждениях кожи и т.д. В целом, миграция обеспечивает доставку клеточного материала в нужную область организма.
Следует отметить, что перемещаться могут как отдельные клетки, так и целые клеточные пласты. Последний вариант характерен для эпителиальных клеток, которые тесно прилегают друг к другу боковыми стенками и подстилаются базальной мембраной. Отростчатые или веретеновидные клетки, погруженные в межклеточное вещество, - мезенхимные клетки - более подвижны, не образуют между собой стойких контактов, вследствие этого они мигрируют одиночно или группами.
В) Сортировка
Механизм сортировки и слипания (адгезии) клеток лежит в основе выделения и объединения клеток одного типа среди всех прочих. В процессе развития клетки «узнают» друг друга и сортируются в зависимости от свойств, т.е. образуют скопления и пласты избирательно, только с определенными клетками. Этот механизм крайне важен при формировании зародышевых листков в ходе гаструляции, образовании структур в органогенезе, осуществлении регенеративных процессов и иммунных реакций в постнатальном развитии.
Следует отметить, что перемещаться могут как отдельные клетки, так и целые клеточные пласты
Г) Запрограммированная гибель
Процесс программированной гибели клеток - апоптоз. В эмбриогенезе он является одним из основных механизмов органогенеза и метаморфоза, способствует достижению характерных для определенного биологического вида черт его морфофункциональной организации. В постнатальном развитии апоптоз обеспечивает гибель клеток на терминальных стадиях дифференцировки (например, эритроцитов), стареющих и поврежденных клеток, уничтожение аутореактивных, т.е. действующих против собственных клеток, клонов лимфоцитов и т.д. Помимо этого на протяжении всего развития механизм программированной клеточной гибели обеспечивает регуляцию численности клеток, что в одних ситуациях обеспечивает стабильное состояние организма, в других - рост, в-третьих - атрофию тканей и органов.
В настоящее время различают два принципиально различных типа клеточной гибели: апоптоз (в переводе с греческого «отпадающий») и некроз.
Некроз представляет собой патологическую форму смерти клеток в результате их острого повреждения. Он характеризуется разрывом цитоплазматической и внутриклеточных мембран, что приводит к разрушению органелл, высвобождению лизосомальных ферментов и выходу содержимого цитоплазмы в межклеточное пространство, при этом часто развивается воспалительный процесс, захватывающий территорию от части клетки до целого органа.
В отличие от некроза, апоптоз - генетически контролируемая клеточная гибель, которая приводит к «аккуратной» разборке и удалению клеток. Он широко распространен и типичен для физиологических условий. Очень важно, что при апоптозе не развивается воспалительный процесс и гибель отдельных клеток или их групп происходит избирательно, без повреждения окружающих здоровых клеток. Выделяют два вида программированной клеточной гибели: апоптоз «изнутри» и апоптоз «по команде».
В первом случае задача процесса - убрать поврежденные клетки. Апоптоз запускается сигналами, возникающими внутри самой клетки при неудовлетворительном ее состоянии.
Второй вариант апоптоза наблюдается во вполне нормальных и жизнеспособных клетках, которые с позиции целого организма оказываются ненужными или вредными. В этом случае клетка получает из внеклеточной среды, например от окружающих клеток, сигнал «погибнуть», который передается через мембранные или, реже, цитоплазматические рецепторы.
Д) Агдезия
Адгезия микроорганизмов (microorganisms adhesion) [лат. adhaesio — прилипание; греч.mikros — маленький и лат. organismus — живое тело, живое существо] — способность микроорганизмов адсорбироваться на твердых поверхностях и чувствительных клетках с последующей колонизацией.
Одним из актуальных направлений современной микробиологии является изучение адгезивного процесса различных микроорганизмов. Известно, что способность микроорганизмов приживаться в ЖКТ, создавая защитный барьер, обусловлена их адгезивными свойствами. Адгезия - это межклеточное взаимодействие, выражающееся в прочном прикреплении клеток к субстрату. Следует отметить, что от адгезивных свойств во многом зависят стабильность и защитные свойства микрофлоры макроорганизма.
Е) Дифференцировка (лабильная и стабильная)
Еще один клеточный механизм развития - дифференцировка клеток. Именно благодаря ей однородный клеточный материал зародыша становится разнородным, образует ткани, входит в состав различных органов и систем, т.е. дифференцировка клеток является основой процесса дифференциации частей и структур зародыша.
Клеточной дифференцировкой (цитодифференцировкой) называется процесс приобретения клетками биохимических, морфологических и функциональных различий. Другими словами, это процесс, в результате которого клетка становится специализированной, имеющей характерное строение, определенный тип метаболизма, и способной к выполнению определенных функций.
Лабильная дифференцировка. Это такое явление, когда пересаженная на другое место часть зародыша не развивается в ту часть, какая должна была быть изначально.
Основные заслуги в изучении этого вопроса и открытие организационных центров принадлежит Г. Шпемену. Он проделал следующие эксперименты. Спинную эктодерму гаструлы зародыша тритона, которая в нормальных условиях превращается в зачаток нервной трубки, пересадил на брюшную сторону зародыша и из этого зачатка развилась кожа живота. В свою очередь на спинную сторону пересадил участок, из которого в нормальных условиях образуется кожа брюшной стороны, то на новом месте этот участок развился в нервную трубку. Из этого эксперимента ясно, что судьба зачатков в стадии ранней гаструлы еще не детерминирована, то есть, не определена, так как участок спинной трубки на брюшной стороне не развился в нервную трубку, когда его пересадили, а превратился в кожу живота, а участок с брюшной стороны, пересаженный на спинную сторону не развился в кожу живота, а превратился в нервную трубку.
Стабильная детерминация или независимая дифференцировка – это такое явление, когда пересаженная на другое место часть зародыша продолжает развиваться в ту часть, какая должна быть независимо от окружения. Например, участок нервной трубки, пересаженный на брюшную сторону в более поздний период, продолжает развиваться в нервную трубку на брюшной стороне. В развитии организма обычно наблюдается переход от зависимой дифференцировки к независимой, т. е. от лабильной детерминации к стабильной.
3. Методы экспериментальной эмбриологии.
Опыты В. Ру и Д. Гердона, Г. Шпемана, Г. Дриша по изучению закономерностей эмбриогенеза.
Методы эмбриологии:
Наблюдения за живыми зародышами с применением кино- и видеосъемки;
Изучения фиксированных срезов;
Маркировки;
Микрохирургии;
Эксплантации;
Трансплантация ядер;
Экстракорпорального оплодотворения.