ВОЗБУДИМЫЕИМЫЕ ТКАНИАНИ
Возбудимыми
являются
нервная и мышечная ткани
ВОЗБУДИМОСТЬДИМОСТЬ -– это свойство клеток
отвечать на действие раздражителя возбуждением (генерацией ПД -– потенциала действия).).
ВОЗБУЖДЕНИЕЖДЕНИЕ -– это ответная реакция
возбудимой клетки на действие раздражителя. .
Характерным проявлением возбуждения является потенциал действия (импульс, , спайк)

•е РАЗДРАЖИТЕЛИЖИТЕЛИ -– факторы
внешней или внутренней среды, , которые вызывают возбуждение клеток (или генерацию ПД) )


Раздражителиздражители: :
•. Адекватные, , неадекватные. .
•». Пороговые, , допороговые, , сверхпороговые. .

МЕМЬРАННЫЕМБРАННЫЕ
ПОТЕНЦИАЛЫЦИАЛЫ
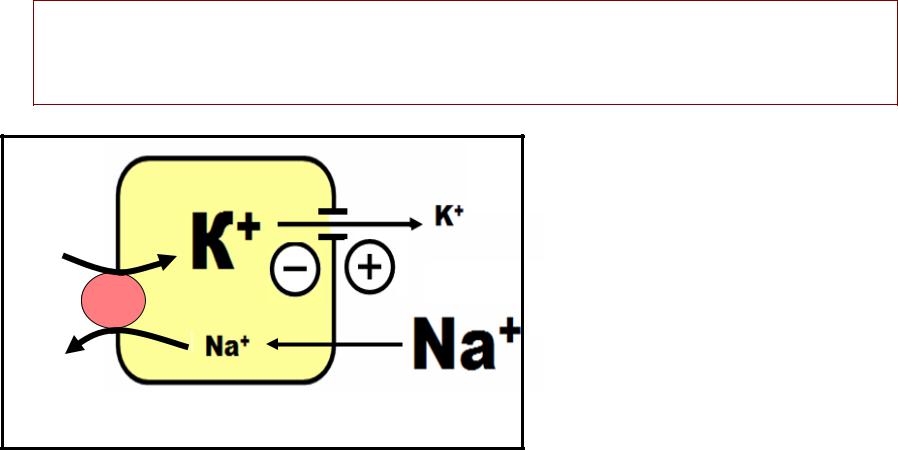
•е ПОТЕНЦИАЛТЕНЦИАЛ ПОКОЯПОКОЯ -– разность потенциалов, , которая существует
между внутренней и наружной поверхностью клеточной мембраны в покое. .
•»е ПОТЕНЦИАЛОТЕНЦИАЛ ДЕЙСТВИЯЕЙСТВИЯ -– быстрое
колебание мембраннного потенциала в ответ на действие раздражителя. .

НЕОБХОДИМЫЕДИМЫЕ УСЛОВИЯЛОВИЯ ДЛЯДЛЯ ФОРМИРОВАНИЯОРМИРОВАНИЯ
ПОТЕНЦИАЛАНЦИАЛА ПОКОЯОКОЯ
•› ИОННАЯ АСИММЕТРИЯ —–
разная концентрация ионов внутри клетки и во внеклеточной среде
•› ПРОНИЦАЕМОСТЬ МЕМБРАНЫ В ПОКОЕ -–
во много раз выше длядля ионовионов калияалия (чем для ионов натрия).).

ВОЗМОЖНЫЕЖНЫЕ ИЗМЕНЕНИЯМЕНЕНИЯ
МЕМЬРАННОГОМБРАННОГО ПОТЕНЦИАЛАНЦИАЛА
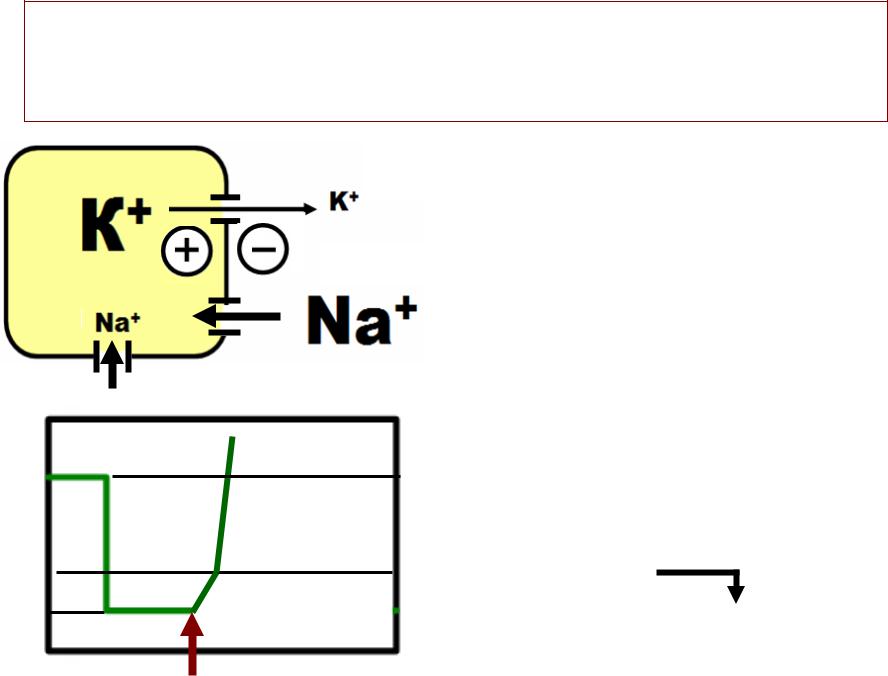
•Мембрана возбудимой клетки в покое поляризована («минус»» внутри и «плюс»» снаружи).). ПП = -70 мВ. .
•ДЕПОЛЯРИЗАЦИЯПОЛЯРИЗАЦИЯ –- это уменьшение внутреннегореннего отрицательного заряда (до 0)..
•ИНВЕРСИЯИНВЕРСИЯ –- это изменение заряда клеточной
мембраны на противоположный (перезарядка до +30 мВ).).
•Деполяризацию и инверсию вызывает вход катионов в клетку (NaМа*,+, Са?+)2+.).
•ГИПЕРПОЛЯРИЗАЦИЯПОЛЯРИЗАЦИЯ –- это увеличение внутреннегореннего отрицательного заряда (до -90 мВ или -100 мВ) )
•Гиперполяризацию вызывает выход катионов из клетки (К+`)) или вход анионов в клетку (ClСТ)-)
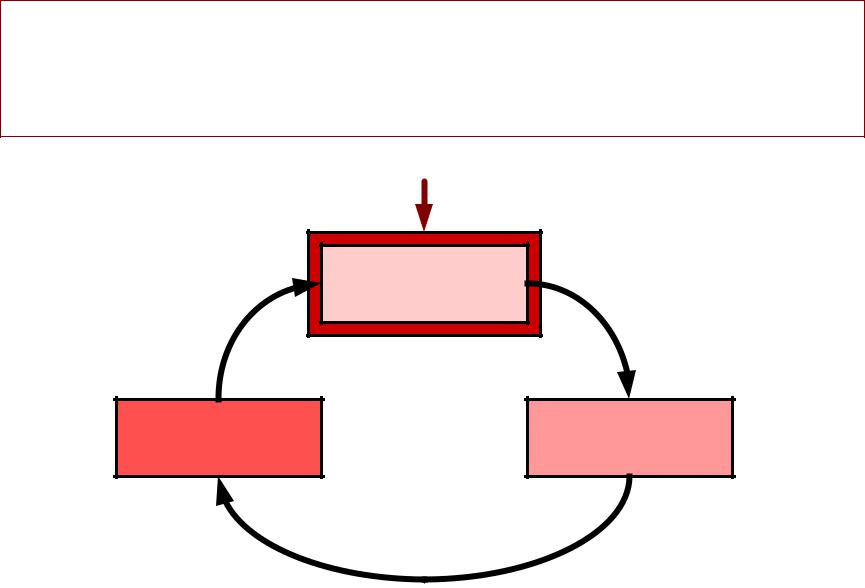
ГЕНЕРАЦИЯАЦИЯ ПДПД: :
циклцикл Ходжкинаоджкина (Hodgkin,Ноадкт, 1958)1958)
Раздражающий стимул
Деполяризация
мембраны
+ +
Вход натрия в клетку
Открывание NaМа - каналов
+
Положительная обратная связь. . Лавинообразное нарастание натриевого тока. . Быстрая деполяризация и инверсия МП. .


ВОССТАНОВИТЕЛЬНЫЙОВИТЕЛЬНЫЙ ПЕРИОДРИОД
(ппослесле генерациирации ПДПД) )
1. Восстанавливается исходный потенциал покоя (реполяризация за счёт движения К+*+ через мембрану) )
2. Восстанавливается исходное состояние ионных каналов (за счётсчет изменения напряженностиряжённости эл..поля) )
3.Восстанавливается исходный градиент концентраций калия и натрия (за счётсчет усиленной работы ионных насосов).).



КРИТЕРИИТЕРИИ ВОЗБУДИМОСТИИМОСТИ
• ВОЗБУДИМОСТЬ -– это способность клетки генерировать импульсы (ПД).).
• Возбудимость нервной ткани выше, , чем возбудимость мышечной ткани. .
• Возбудимость одной и той же клетки может быть повышена, , понижена и даже полностью отсутствовать. .
• Как оценить возбудимость??
АУV -– ПОпоРОГОВЫЙОВЫЙ ПОТЕНЦИАЛНЦИАЛ
Екр |
|
== -5050 |
мВмВ |
|
|
|
|
|
|||
КР |
2020 мВмВ} |
|
|
|
|
|
|
|
|||
|
|
|
|
30 мВ |
|
|
|||||
|
|
|
|
|
|||||||
ЕЕ,0 |
== -7070 мВмВ |
|
|||||||||
|
|
|
|
|
|||||||
|
|
|
|
|
|||||||
|
|
|
|
|
|||||||
Е0 |
= -80 мВ |
||||||||||
|
|
|
|
|
|
||||||
ДУ=VЕ,= Е-0Е–,,Екр
Чем больше пороговый потенциаленциал, , тем меньше возбудимость. .

ПОРОГОРОГ СИЛЫИЛЫ РАЗДРАЖЕНИЯЖЕНИЯ
•‚‹ ПОРОГ СИЛЫ РАЗДРАЖЕНИЯ -– это минимальнаяинимальная сила раздражителя, , которая способна вызвать возбуждение (ПД),),
т..ее.. вызвать деполяризацию мембраны до критического уровня. .
•‚ Раздражители могут быть пороговыми, , допороговыми и сверхпороговыми. .
•‹ Допороговые раздражители возбуждение не вызывают. .
Чем больше порог силы раздражения, , тем меньше возбудимость. .

ПОРОГОРОГ СИЛЫИЛЫ РАЗДРАЖЕНИЯЖЕНИЯ
| | |
| | |
| | |
ПД |
АППИ |
1 |
ПороговыйПороговый
раздражительражитель
раздражающий
стимул
ПОРОГОРОГ ДЛИТЕЛЬНОСТИИТЕЛЬНОСТИ
ДЕЙСТВИЯДЕЙСТВИЯ РАЗДРАЖИТЕЛЯАЖИТЕЛЯ
(ПОРОГРОГ ВРЕМЕНИМЕНИ) )
•‹ ПОРОГ ВРЕМЕНИ —-– минимальныйнимальный промежуток времени, , который необходим для возбуждения клетки под действием раздражителя. .
| \ |
| |
| \ |
Ответная реакция |
| ПОРОГОВЫЙОВЫЙ СТИМУЛИМУЛ
Раздражающий стимул
Чем больше порог времени, ,
тем меньше возбудимость

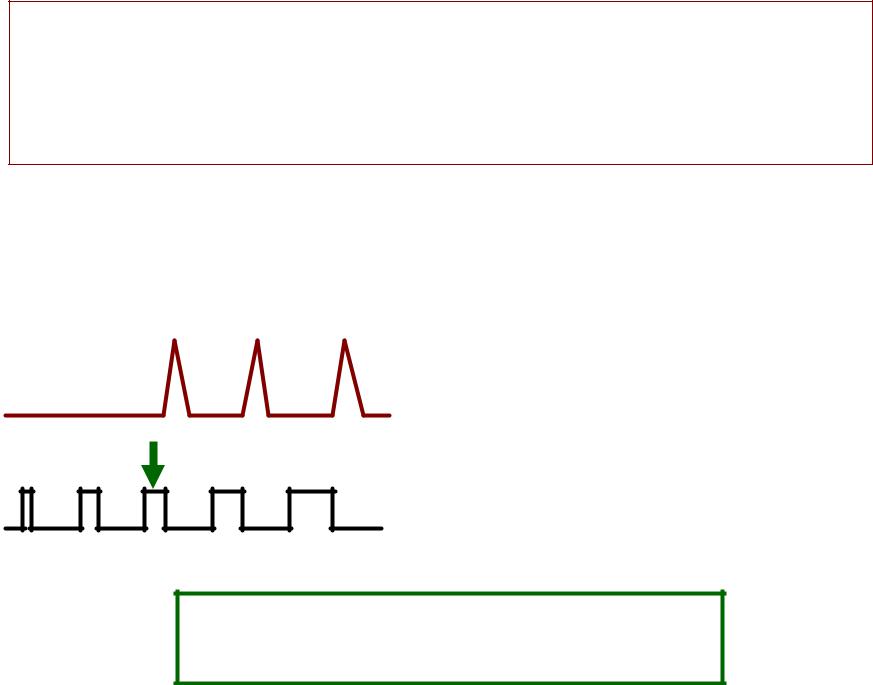
КРИВАЯИВАЯ «СИЛЫЛЫ -– ВРЕМЕНИМЕНИ»»
(ГГоорвегарвега--ВейсаВейса) )
СИЛА (V)
(\)
Р-– реобаза
2рр –- удвоенная
реобаза
пв -– полезное время хр –- хронаксия
2р
р
хр |
пв |
ВРЕМЯ |
|
|
(мсек) |
1.. ЗАВИСИМОСТЬ ОБРАТНАЯ. .
2.. РЕОБАЗАОБАЗА - минимальная
сила раздражителя (постоянного тока),), которая вызывает возбуждение. .
ПОЛЕЗНОЕОЛЕЗНОЕ ВРЕМЯЕМЯ –- время
действия силы, , равной одной реобазе. .
3.. ХРОНАКСИЯОНАКСИЯ -– время действия силы, , равной удвоенной реобазе. .
ЧемЧем длиннеелинн е хронаксияронаксия, , темем меньшеменьше возбудимостьзбудимость. .
4.. Токи УВЧ не вызывают возбуждение. . Применяются в физиотерапии. .

АККОМОДАЦИЯМОДАЦИЯ ВОЗБУДИМОЙИМОЙ ТКАНИАНИ
Екр
ПД
Раздражающий
стимул
ЧемЧем меньшееньше скоростьорость увеличенияличения силылы раздражителядражителя, ,
темем меньшееньше возбудимостьдимость клеткилетки. .
(Происходит смещение критического уровня деполяризации, , увеличивается порог деполяризациии (дельта-\)-V)
Причина: : инактивацияинактивация нанатриевыхиевых каналовканалов. .

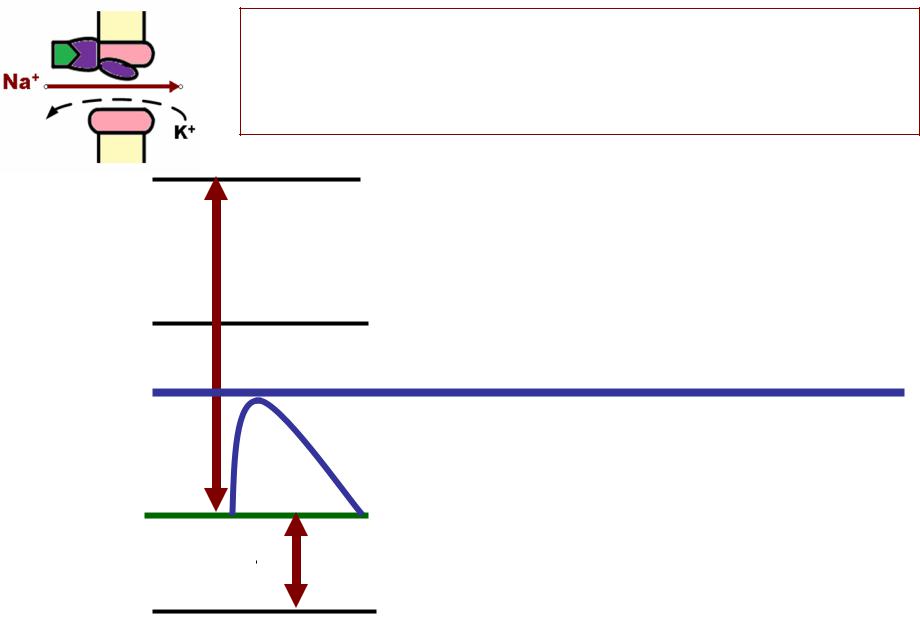
ИЗМЕНЕНИЕМЕНЕНИЕ ВОЗБУДИМОСТИИМОСТИ ВОВО ВРЕМЯЕМЯ ВОЗБУЖДЕНИЯЖДЕНИЯ
1 |
2 3 |
4 |
5 |
ПП
Исходная |
1 |
2 3 |
4 |
5 |
|
|
|
|
|
возбудимость |
|
|
|
|
|
|
|
|
Каждой фазе потенциала действия |
|
|
|
|
соответствует особое состояние |
|
|
|
|
возбудимости |

КАЖДОЙЖДОЙ ФАЗЕФАЗЕ ПОТЕНЦИАЛАНЦИАЛА ДЕЙСТВИЯЙСТВИЯ СООТВЕТСТВУЕТОТВЕТСТВУЕТ ОСОБОЕСОБОЕ СОСТОЯНИЕСТОЯНИЕ
ВОЗБУДИМОСТИДИМОСТИ
ФАЗЫФАЗЫ ВОЗБУЖДЕНИЯЖДЕНИЯ (ПДПД):): |
ФАЗЫФАЗЫ ИЗМЕНЕНИЯМЕНЕНИЯ |
|
ВОЗБУДИМОСТИЗБУДИМОСТИ: : |
1.. Медленная деполяризация |
1.. Супернормальная |
|
возбудимость |
2.. Быстрая деполяризация, , |
2.. Абсолютная рефрактерность |
инверсия |
|
3.. Реполяризация |
3.. Относительная |
|
рефрактерность |
4.. Следовая |
4.. Супернормальная |
деполяризация |
возбудимость |
5.. Следовая |
5.. Субнормальная |
гиперполяризация |
возбудимость |
|
|

СУПЕРНОРМАЛЬНАЯОРМАЛЬНАЯ ВОЗБУДИМОСТЬИМОСТЬ
•‚ Причиной повышеннной возбудимости
является частичная деполяризация клеточной мембраны, , не доходящая до
критическогоического уровня
•. Порог деполяризациии (дельта-\/)-V)
уменьшается
•‚ Клетка при повышеннной возбудимости
способна реагировать не только на пороговые и сверхпороговые, , но даже на
допороговые стимулы
АБСОЛЮТНАЯЛЮТНАЯ РЕФРАКТЕРНОСТЬРАКТЕРНОСТЬ —–
полноелное отсутствиесутствие возбудимостиудимости
•Совпадает с фазой быстрой деполяризациии и инверсиии
•Основной причиной невозбудимости
является инактивацияактивация натриевыхевых каналовналов
•Инактивированные натриевые каналы не могут открываться даже под действием самых сильных раздражителей
•Поэтому клетка во время возбуждения становится невозбудимой
(она не реагирует даже на самые сильные раздражители) )

ОТНОСИТЕЛЬНАЯОСИТЕЛЬНАЯ РЕФРАКТЕРНОСТЬРАКТЕРНОСТЬ
• В процессе быстрой реполяризациии возбуди- - мость клетки восстанавливается от 0 до исходного уровня, , так как
• всё больше натриевых каналов из состояния инактивациии возвращается в исходное состояние (и может быть активировано вновь под действием дополнительного стимула) )
•Относительная рефрактерность -– это
частичная невозбудимость или состояние пониженной возбудимости
•В это время клетка реагирует только на сверхпороговые раздражители
СУБНОРМАЛЬНАЯОРМАЛЬНАЯ ВОЗБУДИМОСТЬДИМОСТЬ
(следоваяследовая) )
•е Следовая гиперполяризация связана с
повышенной проницаемостью клеточной мембраны для калия и выходом ионов К++ из клетки. .
•‚‹ Гиперполяризация приводит к увеличению порога деполяризациии (дельта-\/)-V) и уменьшению возбудимости клетки. .
•» В это время клетка может реагировать только на сверхпороговые раздражители.
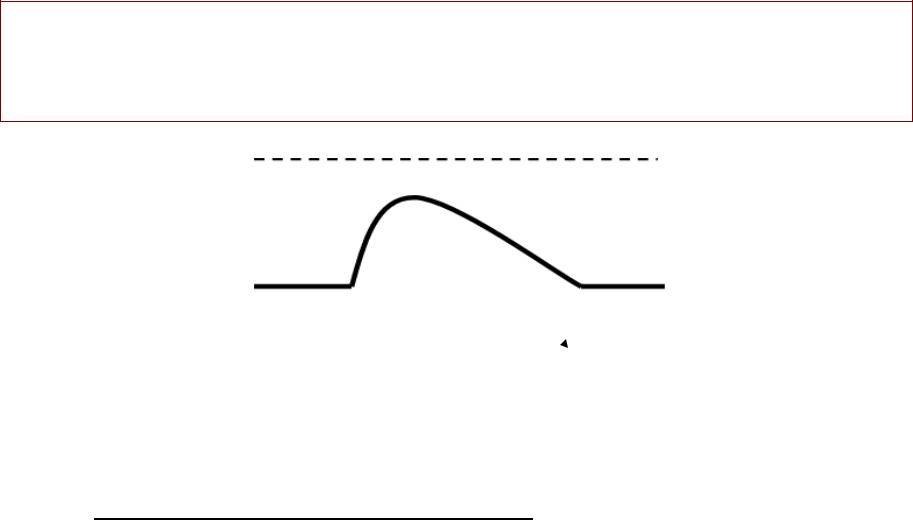
ЛОКАЛЬНЫЙЛЬНЫЙ ОТВЕТТВЕТ -–
реакцияреакция клеткиклетки нана допороговыйдопороговый стимулстимул
Екр = - 50 мВ
Е0 = -70 мВ |
ПП |
|
•.‹ |
Допороговый стимул вызывает{ частичную |
|
|
деполяризацию, , которая не доходит до |
|
|
критического уровня. . |
|
•‚‹ |
Возбуждение не возникает, , т..ее.. генерации ПД не |
|
|
происходит. . |
|
•. |
Свойства локального ответа: : |
|
|
(1) |
не распространяется |
|
(2) |
зависит от силы раздражителя |
|
(3) |
способен к суммациии |
(4) увеличивает возбудимость клетки

СРАВНИТЕЛЬНАЯИТЕЛЬНАЯ ХАРАКТЕРИСТИКАТЕРИСТИКА ПДПД Ии ЛОКАЛЬНОГОАЛЬНОГО ОТВЕТАВЕТА
ЛОКАЛЬНЫЙ ОТВЕТ |
ПОТЕНЦИАЛ ДЕЙСТВИЯ |
||
(1) |
Не распространяется |
(1) |
Распространяется |
(2) |
Зависит от силы раздр. . |
(2) |
Не зависит от силы раздр. . |
(закон силовых отношений) ) |
(закон «всё или ничего»)») |
||
(3) |
Суммируется |
(3) |
Не суммируется |
(4) |
Увеличивает возбудимость| |
(4) |
Во время возбуждения |
|
|
|
возбудимость отсутствует |
|
|
|
(рефрактерность) ) |
|
|
|
|





ВЫВОДЫВОД
• ЛОКАЛЬНЫЙЬНЫЙ ОТВЕТТВЕТ зависит как от силы, , так и от частоты действующих стимулов, , т..ее.. является градуальнымр льным. .
Вот почему в нервной системе он используется для анализа поступающей информации. .
• ПОТЕНЦИАЛТЕНЦИАЛ ДЕЙСТВИЯЙСТВИЯ формируется по закону «всё или ничего»,», имеет постоянную (стандартную) ) амплитуду и форму. . Вот почему в нервной системе импульсы (ПД) ) используются для
кодирования и передачи информациии на большие расстояния..
Лекцияекция 3

НЕРВНЫЕНЫЕ ВОЛОКНАОКНА
Нервное волокно -—–
это отросток нервной клетки


СВОЙСТВАЙСТВА НЕРВНЫХЫХ ВОЛОКОНЛОКОН::
•‚возбудимость
•‚проводимость

ОСНОВНАЯНОВНАЯ ФУНКЦИЯНКЦИЯ НЕРВНЫХЫХ ВОЛОКОНЛОКОН::
•‚Проведение нервных импульсов (ПД)

ЗАКОНЫНЫ ПРОВЕДЕНИЯВЕДЕНИЯ
1.. ЗАКОН АНАТОМИЧЕСКОЙ
И ФИЗИОЛОГИЧЕСКОЙ НЕПРЕРЫВНОСТИ ВОЛОКОН:
<= |
> |
—— |
|
перерезка, , перевязка нервных волокон, ,
действие холода или химических блокаторов прекращает передачу импульсов. .
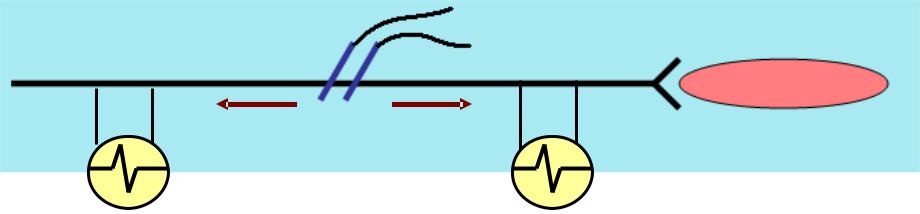
2.. ЗАКОН ДВУХСТОРОНННЕГО ПРОВЕДЕНИЯ: :
стимуляция
по нервным волокнам импульсы проводятся в обе стороны
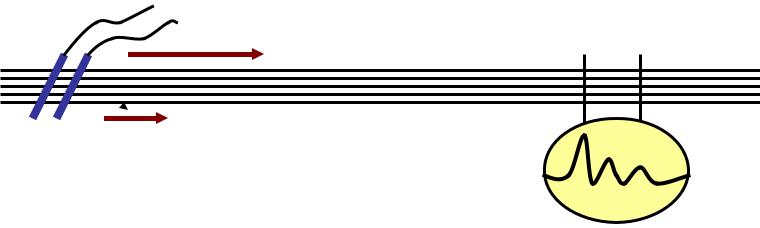
3.. ЗАКОН ИЗОЛИРОВАННОГОНОГО ПРОВЕДЕНИЯ:
пииииииинии >
—
стимуляцияимуляция
А В С
в пучке нервных волокон импульсы не передаются от одного волокна к другому. .

4.. ВЫСОКАЯвысокдя ЛлАБИЛЬНОСТЬ НЕРВНЫХЫХ
ВОЛОКОН
(Лабильность -– это способность клетки генерировать максимальное число ПД
за 1 секунду)
|
|
_ |
|
—я_— |
|
_—_ |
|
|
—_— |
|
|
НЕРВНЕРВ |
СИНАПСИНАПС |
МЫШЦАЫШЦА |
500 имп/сек |
100 имп/сек |
250 имп/сек |
5.. НЕРВНЫЕ ВОЛОКНА ПРАКТИЧЕСКИ НЕ
УТОМЛЯЮТСЯ
Обратимая блокада (ппостоянный ток)
стимуляция ии хо
ОПЫТ ВВЕДЕНСКОГОДЕНСКОГО:
Непрерывная стимуляция нерва продолжалась 8-12 часов. . Каждый раз, , когда блокаду прове- - дения снимали, , импульсы проходили к мышце и мышца сокращалась. .


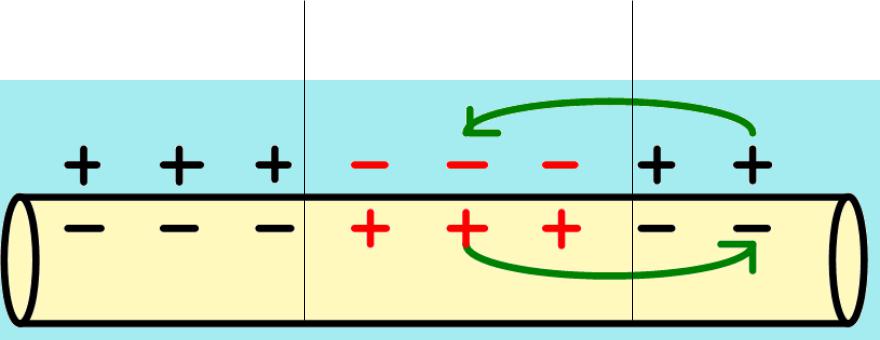
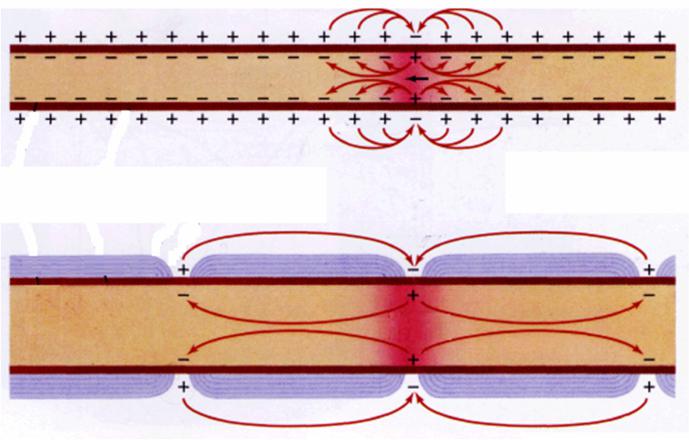
2.. Происходит движение заряженных частиц в электромагнитном поле, , возникают
локальные ионные токи в соответствии с
законом Ома:
[I МИU1 U2
КR

СКОРОСТЬРОСТЬ ПРОВЕДЕНИЯВЕДЕНИЯ ВОЗБУЖДЕНИЯЖДЕНИЯ ПОПО НЕРВНОМУНОМУ ВОЛОКНУЛОКНУ
ЗАВИСИТ
1.. От сопротивления аксоплазмымы (RВ.):):
a
чем больше диаметр волокна, , тем меньше сопротивление аксоплазмымы -–
темм больше скорость проведенияведения!
2.. От сопротивления мембраны (RК):):
м
чем больше сопротивление мембраны, , тем больше
скорость проведения
3.. От амплитуды и длительности ПД:
чем больше амплитуда, , темм больше скорость; чем меньше длительность, , темм больше скорость. .

КАКОЙКОЙ ИМПУЛЬСМПУЛЬС РАСПРОСТРАНЯЕТСЯОСТРАНЯЕТСЯ БЫСТРЕЕЫСТРЕЕ??
А |
Б |
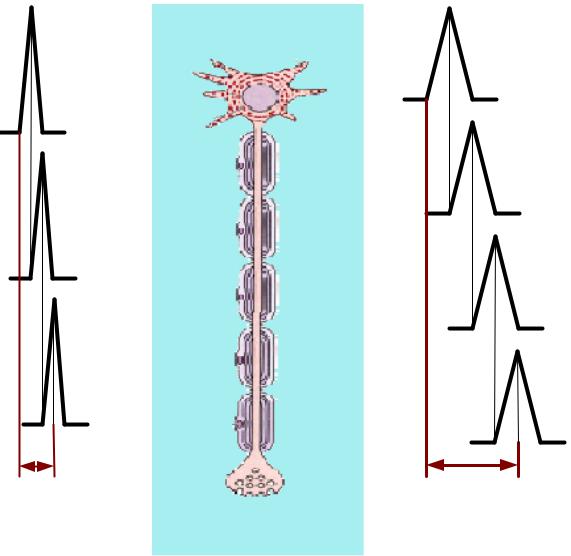



ЗНАЧЕНИЕЧЕНИЕ МИЕЛИНОВОЙЛИНОВОЙ ОБОЛОЧКИОЛОЧКИ
МИЕЛИНИЕЛИН
•. Изолирует нервные волокна. .
•. Выполняет защитную и трофическую функцию. .
•е Увеличивает скорость проведения. .
•. Экономит энергиюию АТФ.

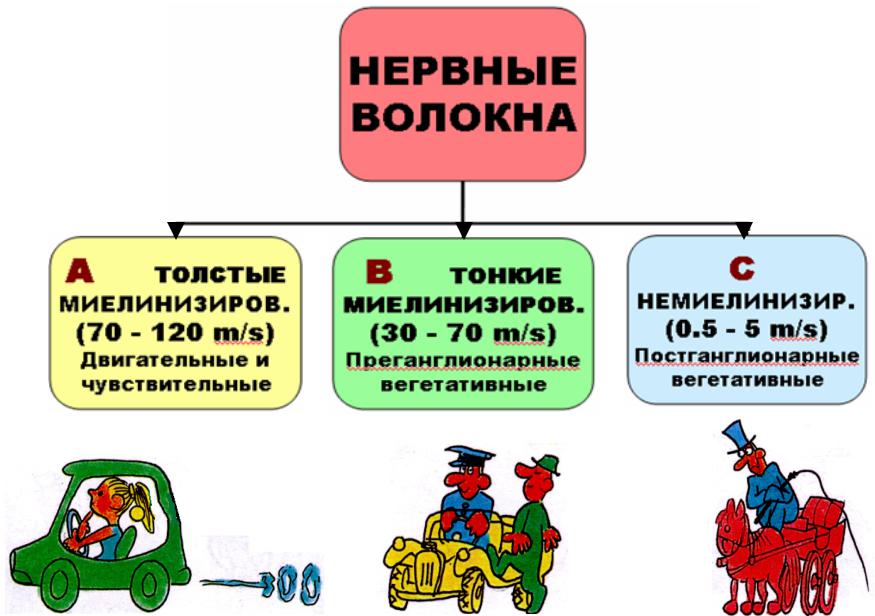

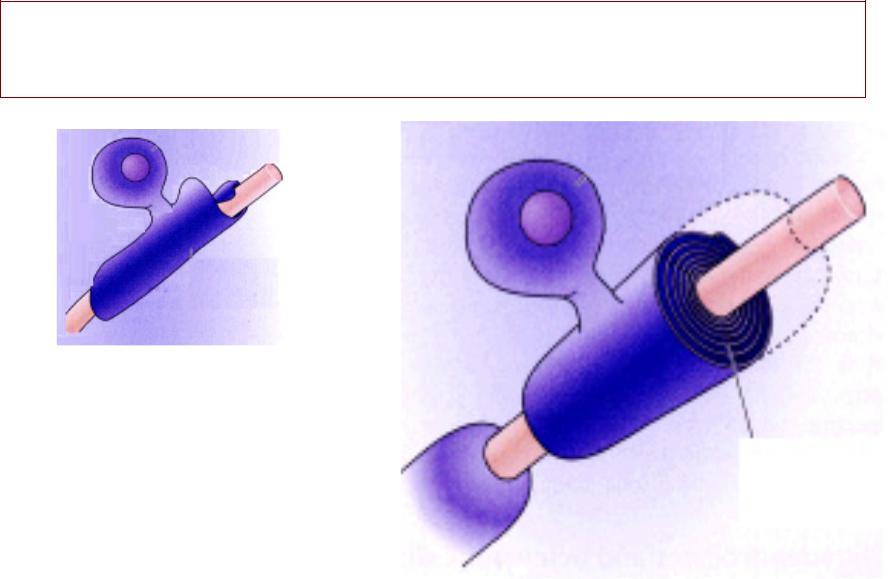
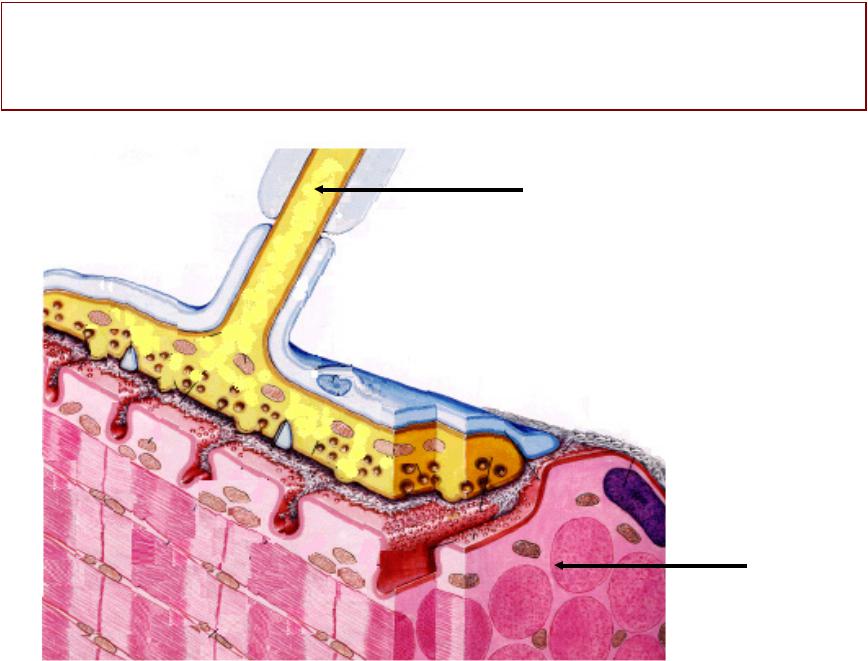
НЕРВНО-ММЫШЕЧНЫЙШЕЧНЫЙ СИНАПСНАПС
Структрура, , с помощью
которой происходит передача возбуждения с нервного волокна на мышечное
МЫШЦА
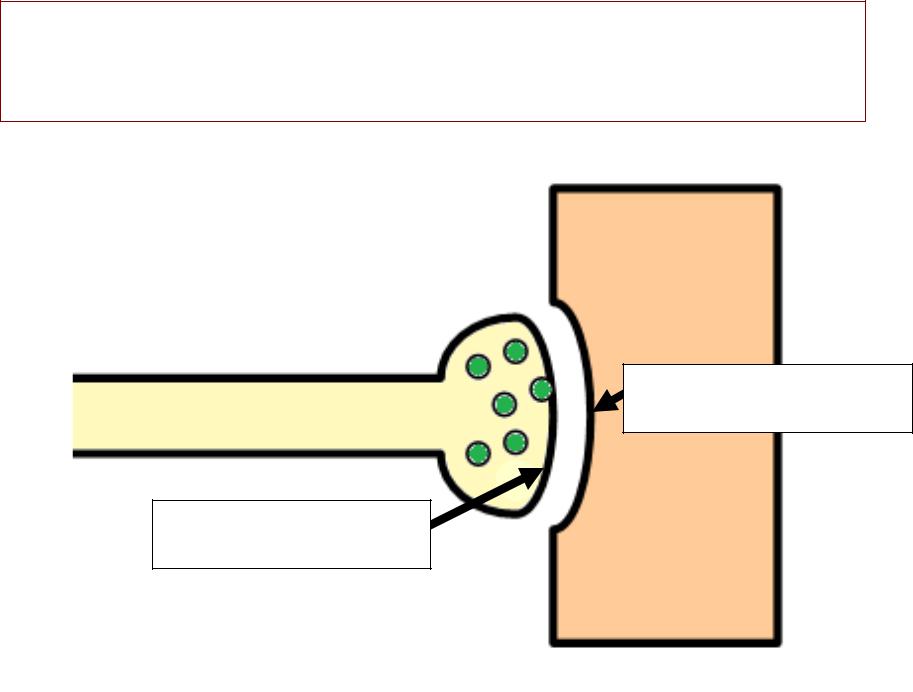

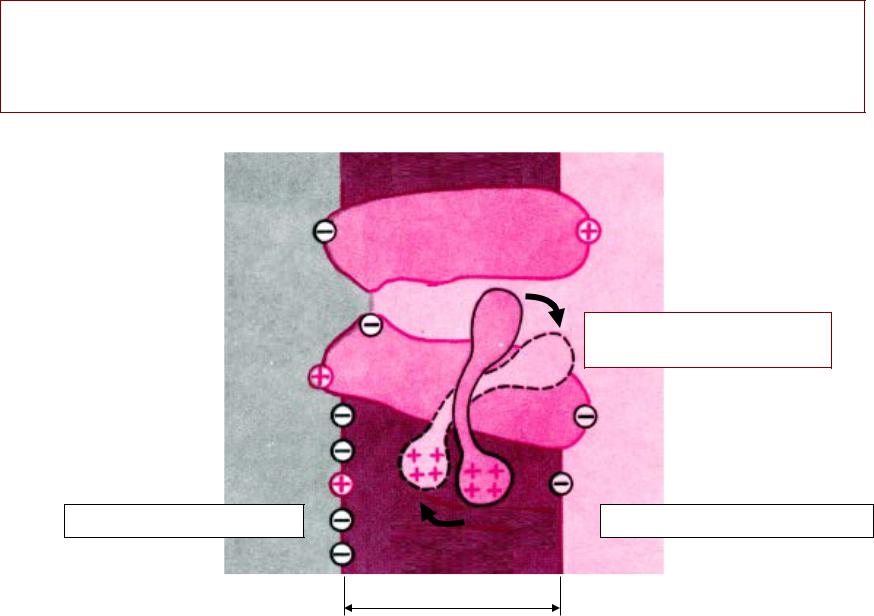
МЕХАНИЗМАНИЗМ ПЕРЕДАЧИДАЧИ ВОЗБУЖДЕНИЯЖДЕНИЯ ВВ СИНАПСЕНАПСЕ
•Механизм химическийимический. .
•Вещество, , с помощью которого передаётся сигнал, , называется «медиатормедиатор»». .
•Медиатором нервно-мышечного синапса является ацетилхолинлхолин (АХ). .
•Ацетилхолине лхолин -– самыймый «быстрыйыстрый»» медиатор:медиатор: уу негоего самыймый короткийоткий медиаторныйиаторный цикликл. .

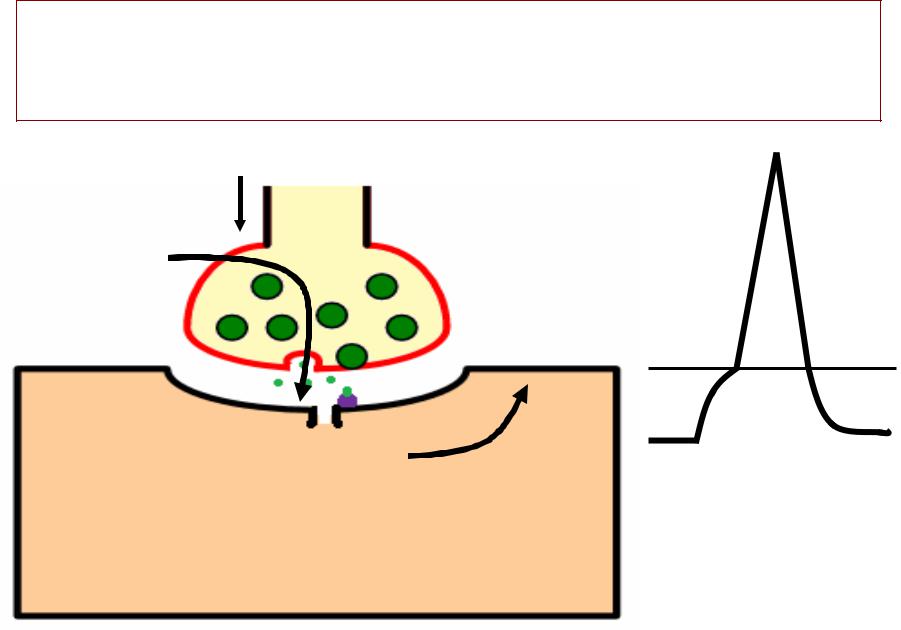
ВЫДЕЛЕНИЕДЕЛЕНИЕ МЕДИАТОРАДИАТОРА
1.. Генерация ПД в нервном окончании
2.. Открытие потенциал-зависимых
кальциевых каналов в мембране нервного окончания и
диффузия ионов кальция (Са) в нервное окончание
3.. Выделение медиатора из синаптиче- - ских пузырьков путёмём экзоцитоза
(оодновременнно из 1000--200 пузырьков)

ДЕЙСТВИЕЙСТВИЕ МЕДИАТОРАДИАТОРА
1.. Взаимодействие медиатора с
рецептором постсинаптической мембраны. .
(Рецепторептор -– этото белковаялковая молекулаолекула, , которая имееет высокое сродство к медиатору. . Рецепторы, , которые связываются с АХ, , называются холинорецепторамиинорецепторами).).
2.. Открытие хемочувствительных ионных каналов постсинаптической мембраны. .
3.. Деполяризация постсинаптической мембраны. . Генерация ПКППКП (потенциала концевой пластинкиинки).
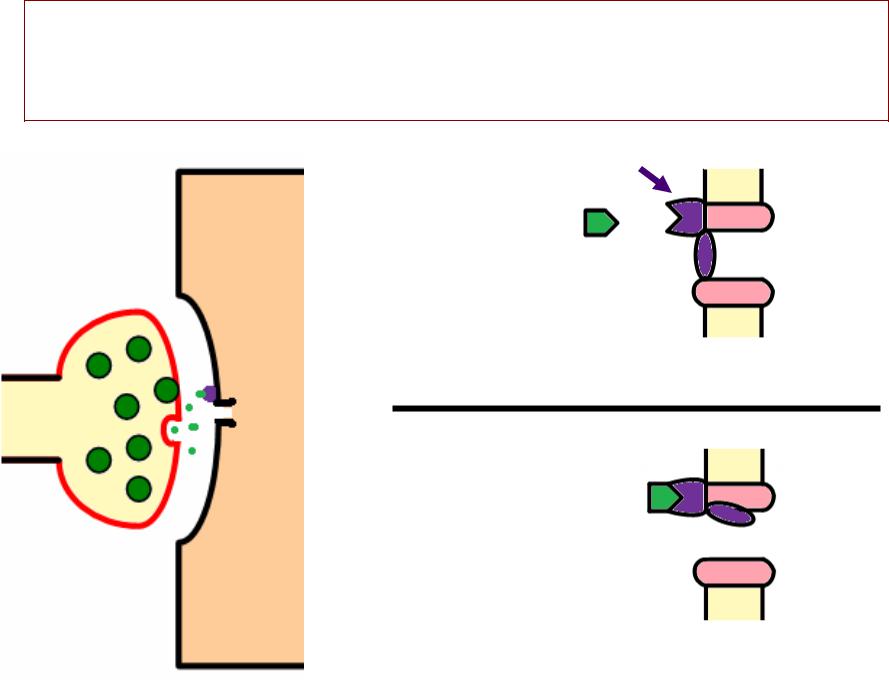

ГЕНЕРАЦИЯАЦИЯ ПКППКП
•› Каналы постсинаптической мембранымбраны:
—– Хемочувствительные
—– Низкой селективности (т..ее.. проницаемы и для натрия, , и для калия)
•»› Ток натрия идетдёт в клетку, , а ток калия в тоо же время идетдёт из клетки
•. Происходит частичная деполяризация
постсинаптической мембраны (ПКП)
•»е ПКП имеет свойства локального ответа


ДАЛЬНЕЙШАЯНЕЙШАЯ СУДЬБАЬБА МЕДИАТОРАДИАТОРА
Самыймый быстрыйыстрый механизмханизм
освобожденияждения рецепторацептора отот медиатораедиатора -—–
ферментативноеферментативное расщеплениещепление
ацетилхолинахолина
АХЭ
Ацетилхолинэстераза
у АЦЕТИЛХОЛИН
2——=>.
ОБРАТНЫЙ
ЗАХВАТ (в нервное
мтокончание)
<— АЦЕТАТ |
ХолинОЛИН |
Метаболизм |
|

ОСОБЕННОСТИБЕННОСТИ ПРОВЕДЕНИЯВЕДЕНИЯ ВОЗБУЖДЕНИЯЖДЕНИЯ ЧЕРЕЗЕРЕЗ СИНАПСНАПС
•‚› Одностороннеее проведение
•‚ Синаптическая задержка
(время, , которое необходимо на проведение возбуждения через синапс = 0..22 мсек)
•.‹ Высокая утомляемость
(связана с истощением запаса медиатора)
•‚‹ Низкая лабильность (100 имп/сек) )
•‚‹ Наличие специфических блокаторов
(например, , ядяд курарераре избирательно блокирует холинорецепторы в нервно-мышечных
синапсах только скелетныхных мышц)

КОНЕЦ

НЕРВНО-ММЫШЕЧНЫЙШЕЧНЫЙ СИНАПСНАПС
Структураруктура, с помощью
которой происходит передача возбуждения с нервного волокна на мышечное

МЕХАНИЗМАНИЗМ ПЕРЕДАЧИДАЧИ ВОЗБУЖДЕНИЯЖДЕНИЯ ВВ СИНАПСЕНАПСЕ
•Механизм химическийимический. .
•Вещество, с помощью которого передаётся сигнал, называется медиаторомедиатором. .
•Медиатор нервно-мышечного синапса -– ацетилхолинлхолин (АХ). .
•Ацетилхолинлхолин -– самый «быстрый»
медиатор: у него самый короткий медиаторный цикл. .
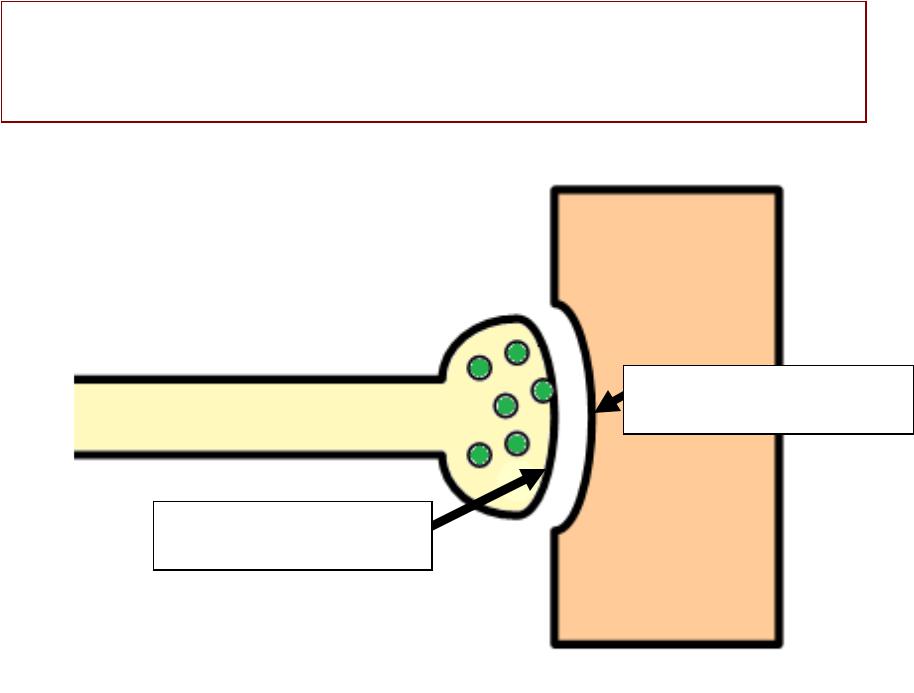

ВЫДЕЛЕНИЕДЕЛЕНИЕ МЕДИАТОРАДИАТОРА
1.. Генерация ПД в нервном окончании
2.. Открытие потенциал-зависимыхх
кальциевыхх каналов в мембране нервного окончания и
диффузия ионов кальция (Са) в нервное окончание
3.. Выделение медиатора из синаптиче- - скихх пузырьков путёмём экзоцитоза
Выделение медиатора одновременно из 1000--200 пузырьков



ЭКЗОЦИТОЗЦИТОЗ

ДЕЙСТВИЕЙСТВИЕ МЕДИАТОРАДИАТОРА
1.. Взаимодействие медиатора с
рецептором постсинаптической мембраны. .
(Рецептор -– это белковая молекула, которая имееет высокое сродство к медиатору. .
Рецепторы, которые связываются с АХ, называются холинорецепторамиинорецепторами).).
2.. Открытие хемочувствительныхх ионныхх каналов постсинаптической мембраны. .
3.. Деполяризация постсинаптической мембраны. . Генерация ПКППКП (потенциала концевой пластинкиинки).
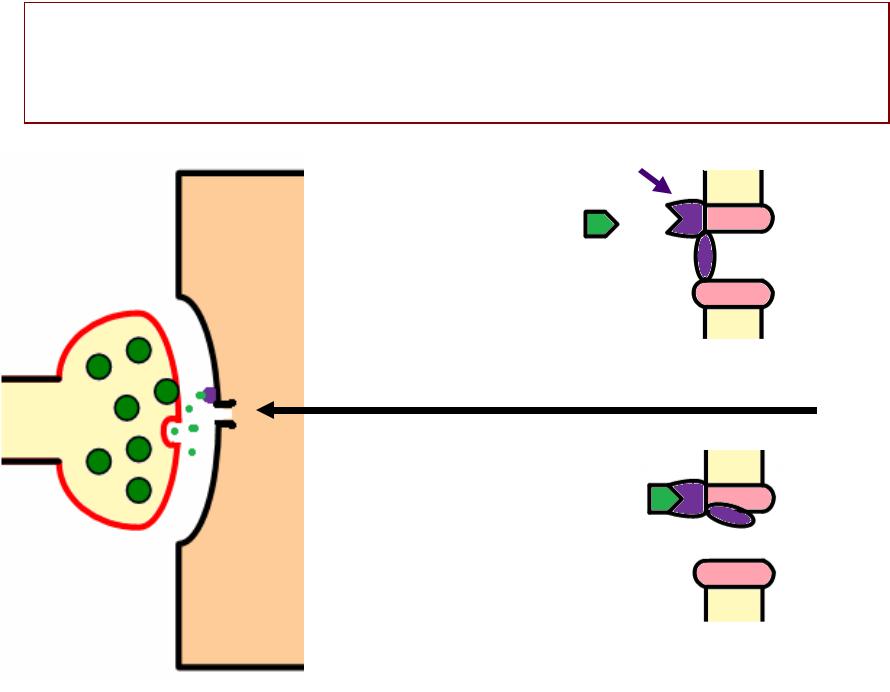

ГЕНЕРАЦИЯАЦИЯ ПКППКП
•› Каналы постсинаптической мембранымбраны:
—– Хемочувствительные
—– Низкой селективности (т..ее.. проницаемы и
для натрия, и для калия)
•»› Ток натрия идетдёт в клетку, а ток калия в тоо же время идетдёт из клетки
•. Происходитходит частичная деполяризация
постсинаптической мембраны (ПКП)
•»е ПКП имеет свойства локального ответа
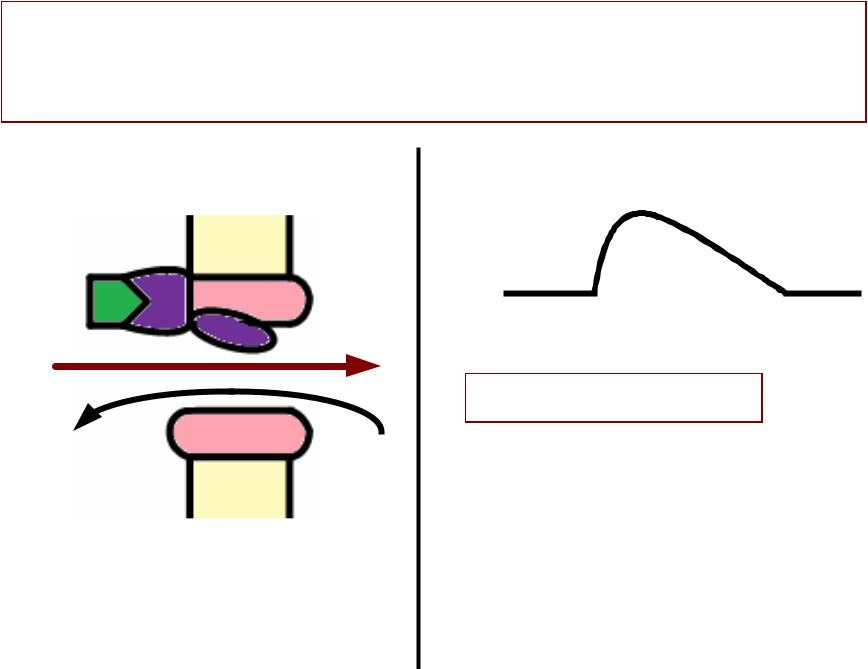
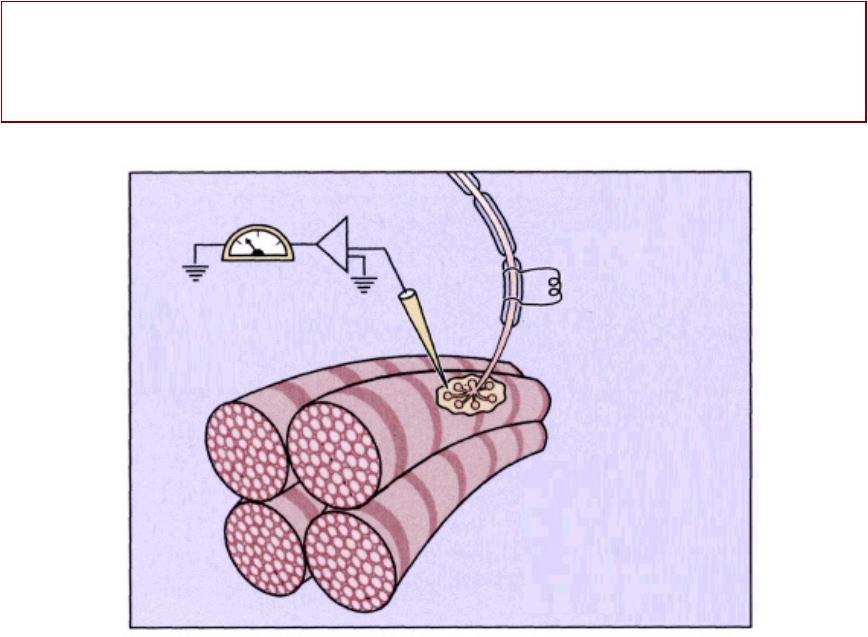
Регистрациястрация ПКПКП сс помощьющью микроэлектродаикроэлектрода
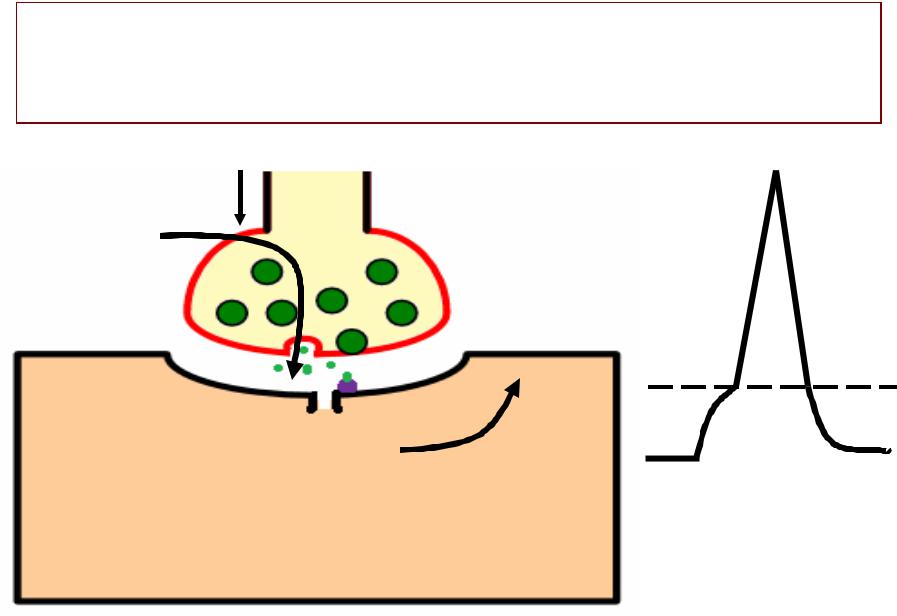

ОСОБЕННОСТИБЕННОСТИ ПРОВЕДЕНИЯВЕДЕНИЯ ВОЗБУЖДЕНИЯЖДЕНИЯ ЧЕРЕЗЕРЕЗ СИНАПСНАПС
•Одностороннеее проведение
•Синаптическая задержка (время,
которое необходимо на проведение возбуждения через синапс -–00.2.2 мсек)
• Высокая утомляемость (связана с
истощением запаса медиатора)
•Низкая лабильность (100 имп/сек)
•Наличие специфическихх блокаторов
(например, ядд кураре избирательно блокирует холинорецепторы в нервно-мышечныхх
синапсахх)
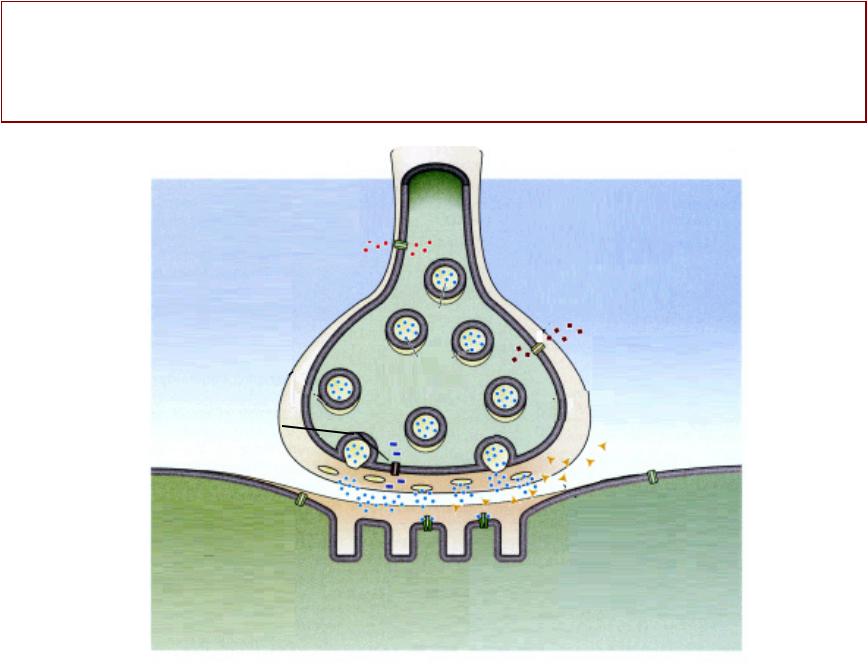
ДАЛЬНЕЙШАЯНЕЙШАЯ СУДЬБАЬБА МЕДИАТОРАДИАТОРА
Самый быстрый механизм освобождения рецептора от медиатора -– ферментативное расщепление ацетилхолина
АХЭ
Ацетилхолинэстеразахолинэстераза
у АЦЕТИЛХОЛИН
2——=>.
ОБРАТНЫЙ
ЗАХВАТ (в нервное
ВИокончание)
<— АЦЕТАТ |
ХолинОЛИН |
Метаболизм |
|

СКЕЛЕТНЫЕНЫЕ МЫШЦЫШЦЫ
ОСНОВНЫЕ ФУНКЦИИ:И:
11..
м2.
3.
ро4.
5.
чех6.
7.
ПОЗАПОЗА -– положение телаа в
пространстве
ЛОКОМОЦИЯМОЦИЯ -– передвижение
телаа в пространстве ТЕПЛОПРОДУКЦИЯПРОДУКЦИЯ
ЗАЩИТНАЯЩИТНАЯ ФУНКЦИЯНКЦИЯ
(например, для органов
брюшной полости)
ДЫХАНИЕЫХАНИЕ КРОВОТОКВОТОК РЕЧЬЧЬ
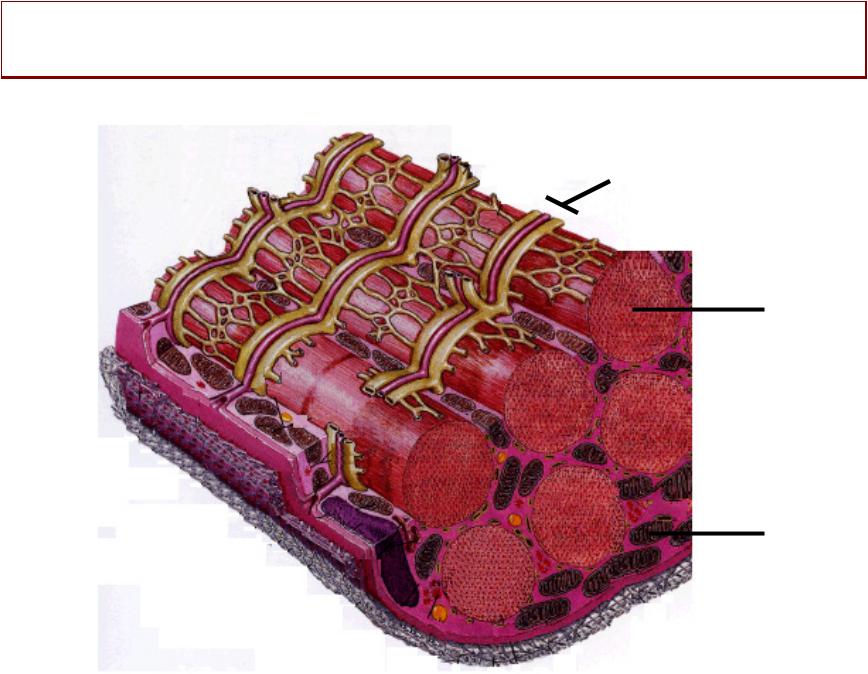
СВОЙСТВАЙСТВА ПОПЕРЕЧНОПОЛОСАТЫХОЛОСАТЫХ МЫШЕЧНЫХШЕЧНЫХ ВОЛОКОНЛОКОН::
•› ВОЗБУДИМОСТЬ
•› ПРОВОДИМОСТЬ
•› СОКРАТИМОСТЬ
•. ЭЛАСТИЧНОСТЬ

МЫШЕЧНОЕЫШЕЧНОЕ ВОЛОКНООКНО -–
СТРУКТУРНАЯ ЕДИНИЦА
СКЕЛЕТНОЙ МЫШЦЫ
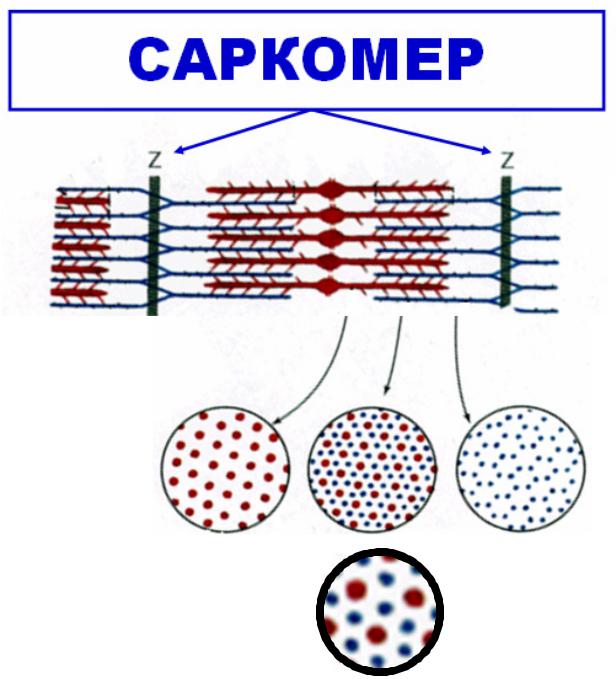
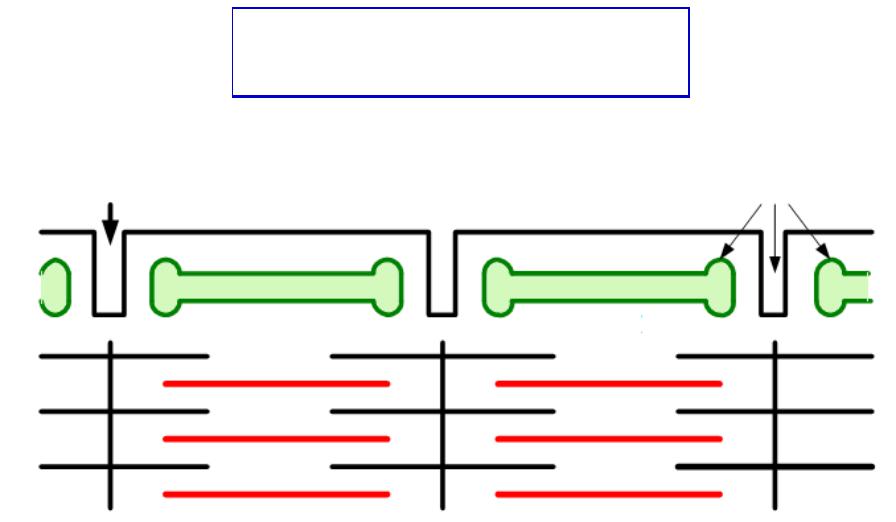
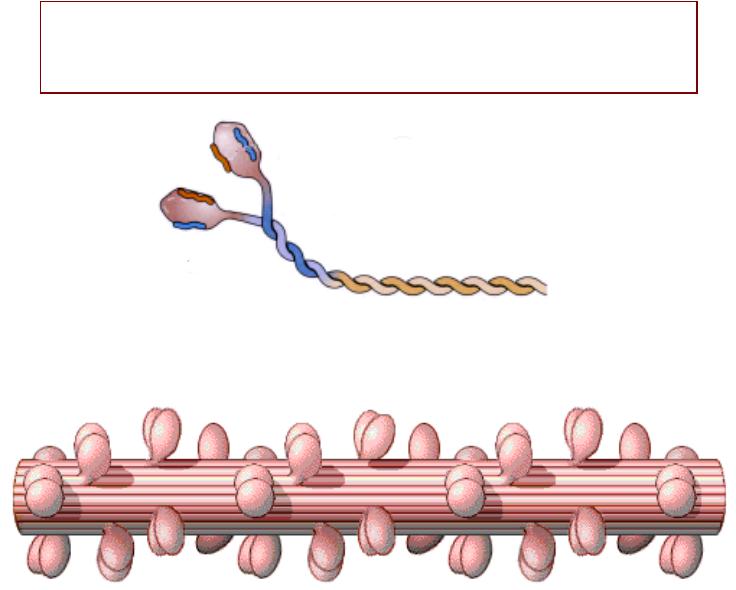

МЕХАНИЗМАНИЗМ СОКРАЩЕНИЯЩЕНИЯ
•Возбуждение мышечного волокнаа
•Распространение ПД по мембране (в том числе ив в Т-трубочкахах)
•Выделение ионов СаСа?*2+ из саркоплазматиати- - ческого ретикулумаа (СПР)
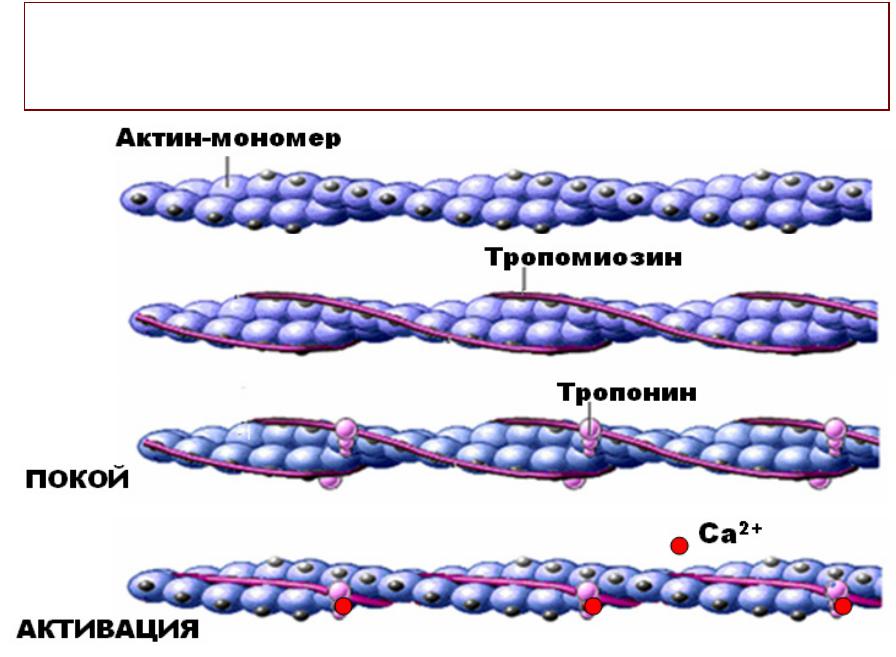
•Взаимодействие СаСа?*2+ с тропонином
•Активацияация актиновых нитей
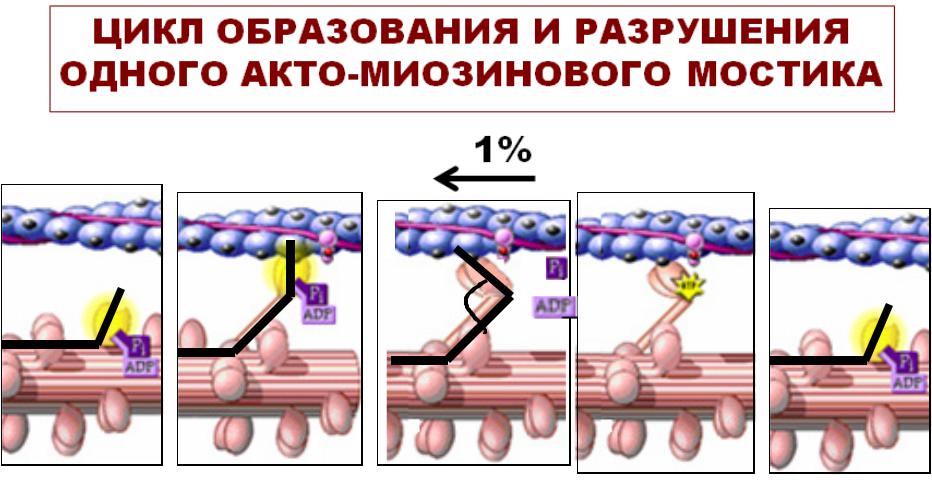
•Образованиеание акто-миозиновых мостиков
•Скольжение нитей актинаа и миозинаа
•Укорочение каждого саркомераа
•Укорочение (сокращение) мышцы в целом. .


МЕХАНИЗМАНИЗМ РАССЛАБЛЕНИЯАБЛЕНИЯ
•Активацияивация СаСа?*2+- - насосасоса в мембранеане СПР
•Активный транспорт СаСа?+2+ из саркоплазмыазмы
обратноатно в СПР
•Возвращение актинаа в состояние покоя
•Прекращение взаимодействия актинаа и миозинаа
•Возвращение каждого саркомераа к
исходной длине (заа счёт эластическихических свойств скелетной мышцы)
•Расслабление мышцы. .
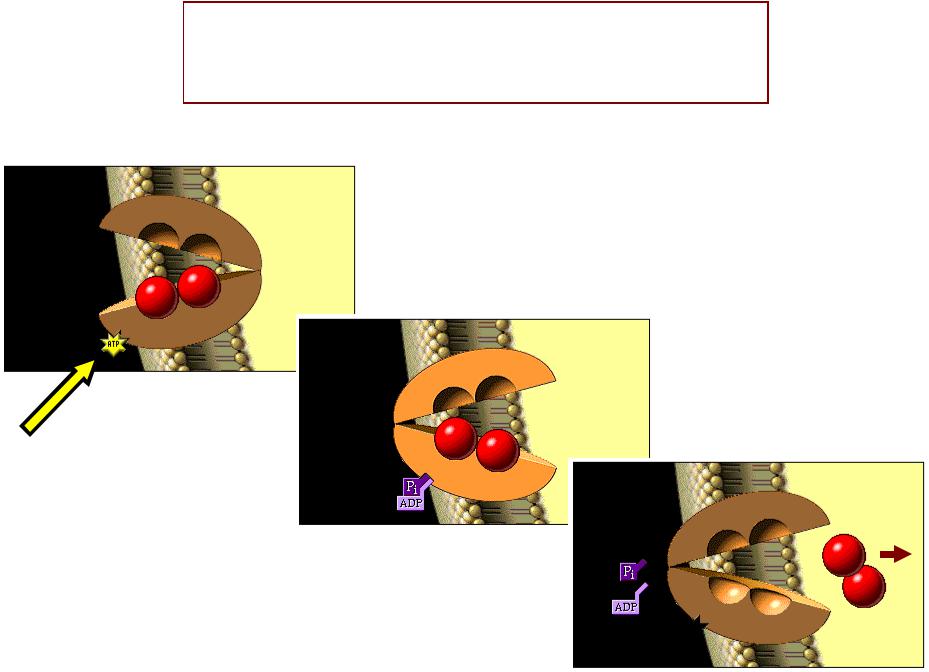
СИЛАИЛА СОКРАЩЕНИЙЩЕНИЙ ОДИНОЧНОГОНОЧНОГО МЫШЕЧНОГОЫШЕЧНОГО ВОЛОКНАЛОКНА ЗАВИСИТИСИТ ОТОТ КОЛИЧЕСТВАИЧЕСТВА АКТО-МИОЗИНОВЫХОВЫХ МОСТИКОВОСТИКОВ::
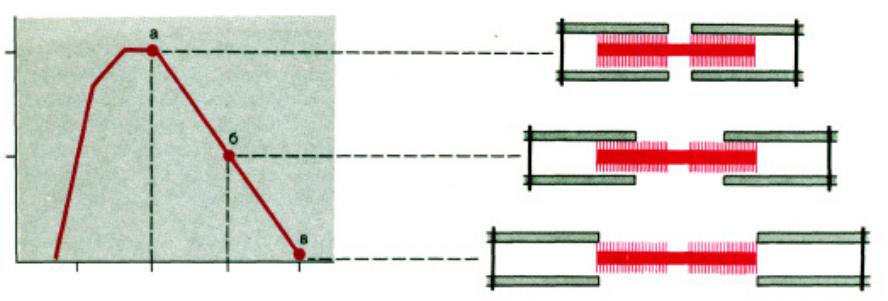
•е Чем больше концентрация ионов Саа?2+ в саркоплазмеазме, темм больше силаа сокращенияащения
(например, при ритмической стимуляциии мышцы и
формированиии тетанусаа)
•‚ Чем больше исходнаяходная длинаина саркомераркомера, , темм больше силаа сокращения (закон Франка-а-
Старлингаа)
•.‹ Гипертрофияфия мышц увеличивает силу
сокращения (так как увеличивается количество актинаа, миозина,а, миофибриллл в каждом мышечном
волокнене)
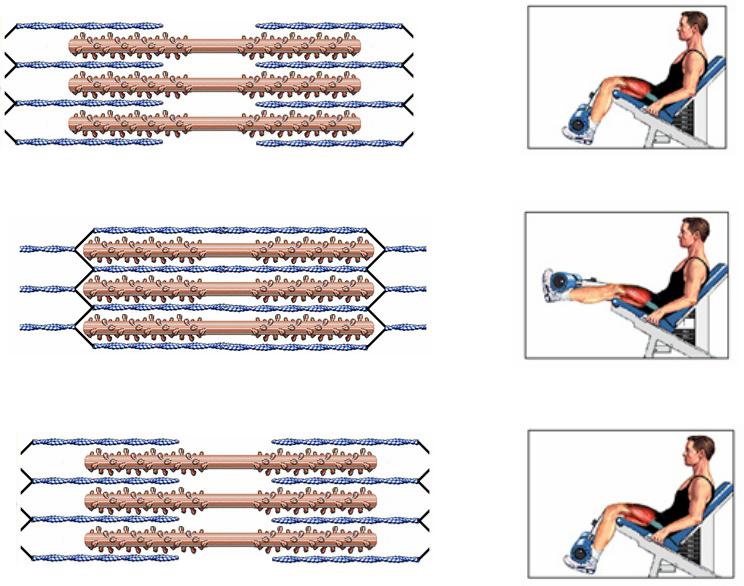
СОКРАЩЕНИЕЩЕНИЕ ИЗОЛИРОВАННОЙОВАННОЙ СКЕЛЕТНОЙЕТНОЙ МЫШЦЫЫ
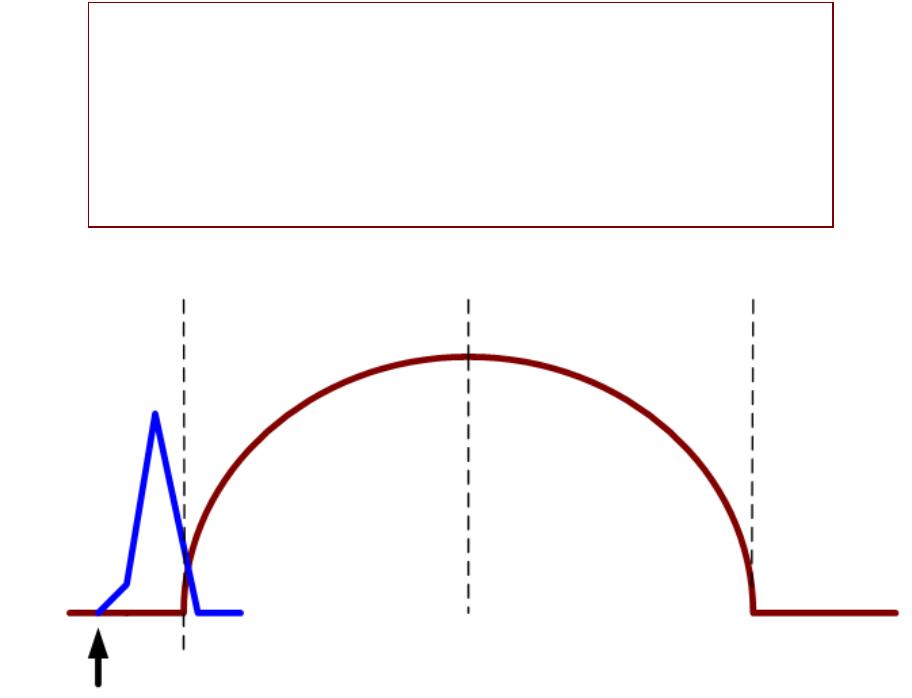
Парныерные стимулыулы вызываютывают суммациюмацию мышечныхшечных сокращенийщений
НЕПОЛНАЯОЛНАЯ СУММАЦИЯМАЦИЯ::
второй стимул поступает к мышце в фазу расслабления
ПДПД |
APПД |
AP |
|
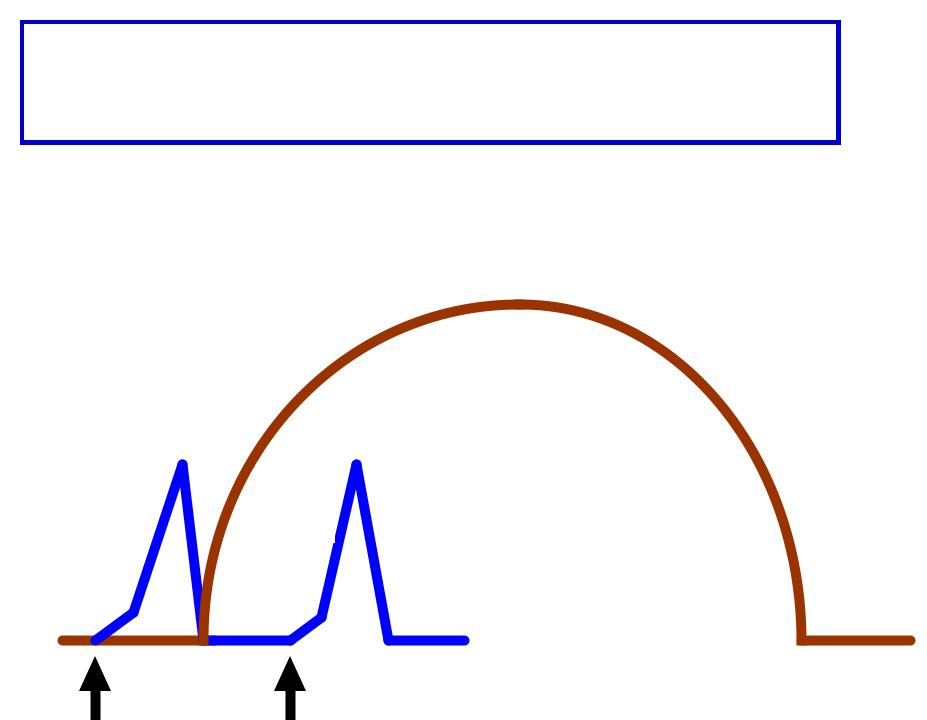
Парныерные стимулыулы вызываютывают суммациюмацию мышечныхшечных сокращенийщений
ПОЛНАЯЛНАЯ СУММАЦИЯМАЦИЯ::
второй стимул поступает к мышце в фазуазу сокращения
AP_ ПДПД APПДпд

Ритмическаямическая стимуляциямуляция вызываетзывает формированиеформирование тетанусануса
ЗУБЧАТЫЙЫЙ ТЕТАНУСАНУС -–
Результатат неполной суммации,и, когдаа
каждый последующий стимул поступает к мышце в фазуазу расслабленияабления
APПД
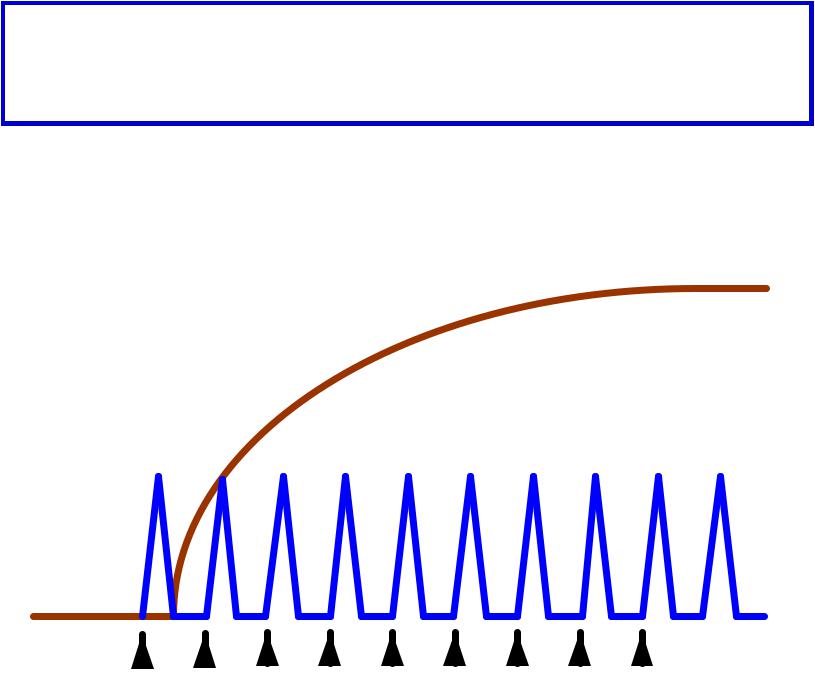
Ритмическаяическая стимуляциямуляция вызываетзывает формированиеформирование тетанусануса
ГЛАДКИЙДКИЙ ТЕТАНУСАНУС —–
результатат полной суммацииации, когдаа каждыйаждый последующий стимул поступает к мышце в фазу сокращения
APПД„
|/
АКАКАХЁЕЁЕЁА

ДВИГАТЕЛЬНАЯИГАТЕЛЬНАЯ ЕДИНИЦАНИЦА
Функциональной единицей скелетныхных мышц является двигательнаяельная (нейроо-моторнаяая) единица,а, в составав которой входит:
•.‹ Альфаа--мотонейрон спинного мозгаа,
•. аксон мотонейронаа
•. и все мышечные волокна,а, которыеые им иннервируются. .
аксон

ДВИГАТЕЛЬНЫЕЛЬНЫЕ ЕДИНИЦЫИЦЫ
В составе двигательной единицы
может быть разное количество мышечных волокон (в разных мышцахах):
1--22 мышечных волокнаа __вв мышцахах гортаниани
5--77 мышечных волокон____в глазодвигательных м.. 10--15 мышечных волоконвв мышцахах пальцев рук
200--2000 волокон__________ в больших мышцахах ног, спины и т..пп.. (ттонус!)
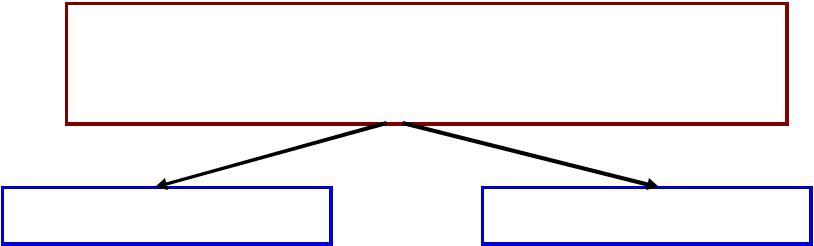
КЛАССИФИКАЦИЯФИКАЦИЯ ДВИГАТЕЛЬНЫХНЫХ ЕДИНИЦНИЦ
|
м |
|
||||
|
МЕДЛЕННЫЕЛЕННЫЕ |
|
|
БЫСТРЫЕЫСТРЫЕ |
||
Свойстваа мышечных |
|
Свойстваа мышечных |
||||
|
волокон: |
|
|
|
волокон: |
|
красные |
|
белые |
||||
слабые |
|
сильные |
||||
аэробные |
|
анаэробные |
||||
содержатат миоглобин-О-О. | |
содержатат креатинфосфатат |
|||||
триглицериды |
2 |
гликоген |
||||
|
||||||
не утомляются |
|
быстро утомляются |
||||
выполняют тоническую| |
выполняют локомоторную |
|||||
|
функцию |
|
функцию |
|||
|
|
|
|
|
|
|



ФИЗИОЛОГИЯИЗИОЛОГИЯ ГЛАДКИХДКИХ МЫШЦЫШЦ
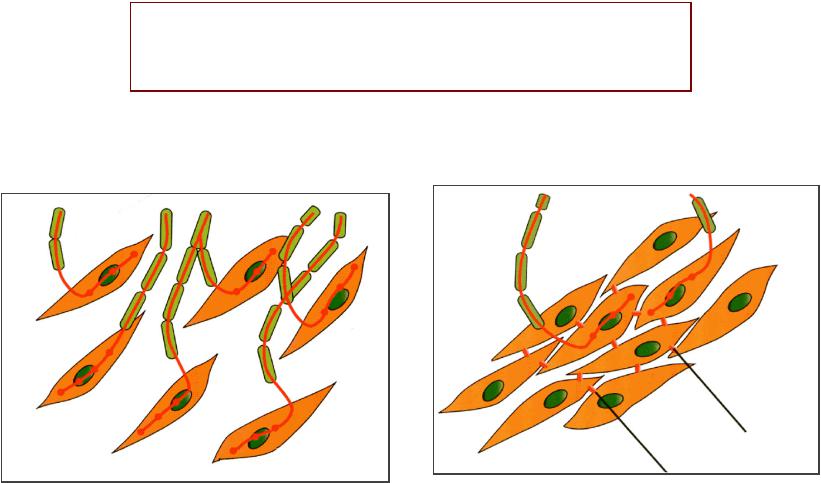
ГЛАДКОМЫШЕЧНЫЕШЕЧНЫЕ ВОЛОКНАОКНА (оособенностинности структурыуктуры):):
•Веретенообразныеые клетки
•Длиной до 500 мкм
•Без поперечной исчерченности
•С одним ядром
•Способныы к делению
•Содержат актин и миозин
•Сокращаются с помощью механизма
«скользящих нитей»

ОСОБЕННОСТИБЕННОСТИ ВОЗБУЖДЕНИЯЖДЕНИЯ

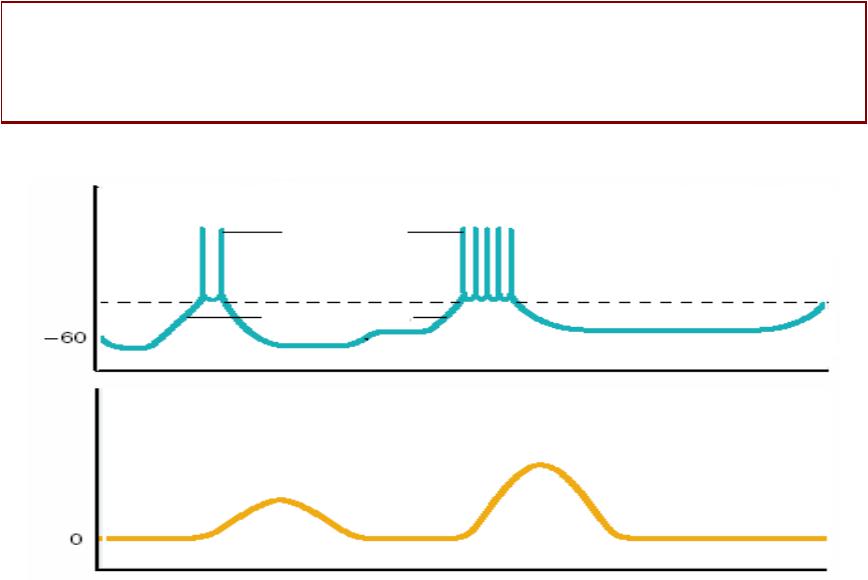
МЕДЛЕННЫЕЛЕННЫЕ ВОЛНЫНЫ ДЕПОЛЯРИЗАЦИИЛЯРИЗАЦИИ
•ПОТЕНЦИАЛНЦИАЛ ПОКОЯОКОЯ низкий –- примерно минус 500--60 мВ, непостоянныйый. .
•МЕДЛЕННЫЕЛЕННЫЕ ВОЛНЫЛНЫ ДЕПОЛЯРИЗАЦИИОЛЯРИЗАЦИИ
формируются за счёт попеременнного участия потенциалчувствительныхльных Са-каналов (вход Са и деполяризация) и кальций-зависимыхых К-каналов (ввыход калия и гиперполяризация). .
Колебания МП происходят на фоне постоянной работыы ионныхых насосов. .
Частота: 3-15 волн в мин; Амплитуда: 10--20 мВ
• МЕДЛЕННЫЕ ВОЛНЫ сопровождаются усилением тонуса гладких мышц (во время деполяризации)и) и ослаблением тонуса (во время гиперполяризации). .
(ММИОГЕННЫЙ ТОНУС –- это постоянное сокращение гладкооммышечныхых клеток без участия нервной системыы). .

АВТОМАТИЯМАТИЯ
• АВТОМАТИЯ –- это способность
гладкокоммышечныхых клеток самостоятельно генерировать ПД без участия нервныхых или гуморальныхых раздражителей. .
•На вершине волныы медленной деполяризациии спонтанно возникают потенциалыы действия. . Критический
уровень для возбуждения висцеральныхых гладких мышц примерно минус 35 мВ. .
•Для возникновения ПД медленная деполяризация должна произойти одновременнно в 30--40 гладкомышечныхых клетках. .


ОСОБЕННОСТИБЕННОСТИ ПРОВЕДЕНИЯВЕДЕНИЯ
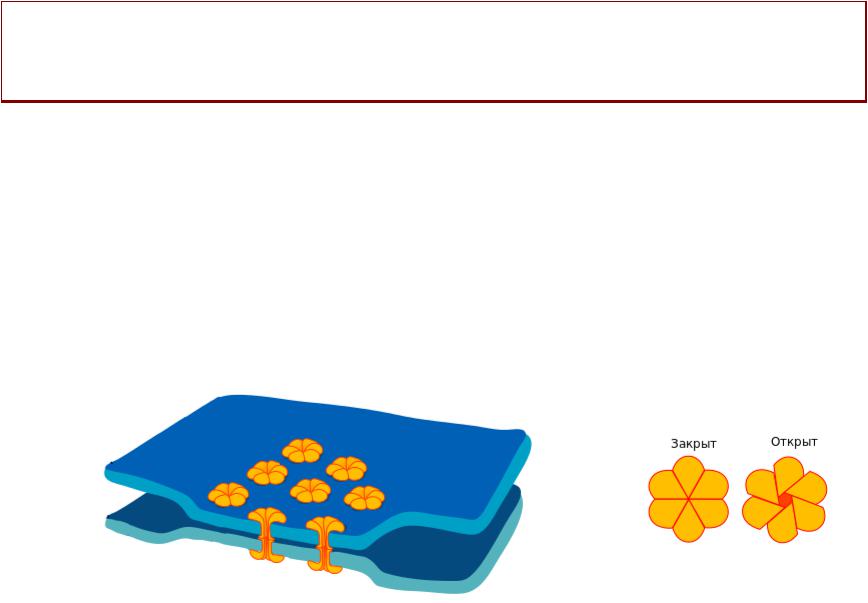
ФУНКЦИОНАЛЬНЫЙНАЛЬНЫЙ СИНЦИТИЙЦИТИЙ
•Пласт гладкооммышечныхых клеток, заключенныхых в
общую фиброзную оболочку, функционирует как единое целое. .
•ПД распространяется от одной мышечной клетки к другой с помощью электрических синапсов
(нексус, щелевой контакт). .
Закрыт Открыт
• Скорость распространения в среднем 2 см/сек. .

ОСОБЕННОСТИБЕННОСТИ СОКРАЩЕНИЯЩЕНИЯ


ОБРАЗОВАНИЕОВАНИЕ АКТО-МИОЗИНОВЫХОВЫХ МОСТИКОВОСТИКОВ
•Когда лёгкая цепь миозина не фосфорилирована, циклического прикрепления и отделения миозиновой головки по отношению к актиновой нити не происходит. .
•При фосфорилированиии этой регуляторной цепочки головка миозина приобретает способность к осуществлению всего циклического процесса образования и разрушения акто-миозиновыхых мостиков (как и в скелетной мышце). .
•Таким образом, энергия АТФ в процесссе сокращения затрачивается (1) на фосфорилирование лёгких
цепей миозина и (2) на осуществление цикла «образования -– разрушения» актомиозиновыхых мостиков. .
•(Кроме того, энергия затрачивается на работу KК--МаNa- - насосов, а также Са-насосовов)
АКТИВАЦИЯВАЦИЯ МИОЗИНАИОЗИНА
ВХОД КАЛЬЦИЯ В КЛЕТКУ
+
КАЛЬЦИЙ + КАЛЬМОДУЛИН
АКТИВАЦИЯ ФЕРМЕНТА
киназаназа лёгкихёгких цепейпей миозинаиозина
+
ФОСФОРИЛИРОВАНИЕ МИОЗИНА
у
ОБРАЗОВАНИЕ АКТОМИОЗИНОВЫХ МОСТИКОВ
СОКРАЩЕНИЕЩЕНИЕ ГЛАДКОМЫШЕЧНОГОЫШЕЧНОГО ВОЛОКНАЛОКНА
ДляДля полноголного расслаблениялабления необходимоходимо нене тольколько (1)1) удалениеление кальцияльция изиз саркоплазмыплазмы,,
ноно и и (2)2) дефосфорилированиефосфорилирование лёгкихёгких цепейцепей миозинаиозина сс помощьющью ферментафермента
фосфатазыы лёгких цепей,
активностьвность которойорой нене зависитвисит отот кальцияльция

ФАКТОРЫАКТОРЫ,, ВЫЗЫВАЮЩИЕВ ЮЩИЕ СОКРАЩЕНИЕЩЕНИЕ ГЛАДКИХДКИХ МЫШЦЫШЦ
•.‹ Автоматия пейсмекерныхых клеток
•е Медиаторыы вегетативных нервов
•. Гормоныы
•е Местныеые (тканевыеые) химические
раздражителиражители:
паракринныеые в-ва,
изменение рН,рн осмотического давления, ионного состава и др. .
•е Растяжение

ОДИНОЧНОЕНОЧНОЕ СОКРАЩЕНИЕЩЕНИЕ
• Общеее время сокращения достигает 1--33 сек (что в 30 раз больше, чем у скелетной мышцыы):
•Длинныйый латентныйный период (50-100 мсек)
•Сокращение развивается медленнно. . Проходит 0,5 сек до достижения максимальной силыы сокращения. .
•Расслабление продолжается 1--22 сек. .
•АТФ-азная- активность миозина низкая. . Цикл образования и разрушения акто-миозинового
мостика длиннеее в 10-300 раз. .
•Сила сокращения 3--44 кгг/см?,2, как у скелетныхых
мышц (и даже больше), а энергии на сокращение тратится в 10-300 раз меньше. .
•Гладкий тетанус возникает при низкой частоте
(11 гцц)..

ПЛАСТИЧНОСТЬИЧНОСТЬ
•‚‹ Свойством гладких мышц является
ПЛАСТИЧНОСТЬ: Гладкие мышцыы могут полностью расслабляться и в укороченнном, и в растянутомянутом состоянии.и.

РЕЛАКСАЦИЯСАЦИЯ НАПРЯЖЕНИЯЖЕНИЯ
РЕЛАКСАЦИЯСАЦИЯ НАПРЯЖЕНИЯЖЕНИЯ
•Увеличение объёма полого органа вызывает растяжение мышечной стенкиенки, увеличение силыы её сокращения -– и давление в полости
органа повышается. . Однако через 15--60 сек
сила сокращения и давление внутри полого органа возвращаются к исходному уровнюню
(до следующего повышения объема и ещё большего растяжения стенкиенки). .
ОБРАТНАЯРАТНАЯ РЕЛАКСАЦИЯСАЦИЯ НАПРЯЖЕНИЯЖЕНИЯ
•Внезапное уменьшение объёма вызываетывает падение давления в полости органа. . Однако через 15--60 сек сила сокращения мышечной
стенки увеличивается - и давление возвращается к исходному уровню. .
ЗНАЧЕНИЕЧЕНИЕ этихих явлений состоит в том, что (за
исключением коротких периодов времени) они позволяют полому органу поддерживать в его просвете примерно одно и тоо же давлениевление,
несмотря на длительныеые, значительныеые по величине изменения объема. .
МЕХАНИЗМАНИЗМ «ЗАЩЁЛКИЩЁЛКИ»»
•Механизм «защёлки» необходим для длительного поддержания тонуса гладких мышц (без больших затрат энергии)и). .
•Сразу после развития необходимой силыы сокращения возбуждение гладких мышц снижается до исходного уровня. . При этомом мышца сохраняет
полную силу своего сокращения в течение нескольких часов, потребляяя очень мало энергии (в 300 раз меньше, чем скелетная мышца). .
•Одно из объяснений механизма «защёлки»: когда миозинкиназа и миозинфосфатаза сильно
активированыы, частота циклов миозиновыхых головок
и скорость сокращения высокие. . Затем, когда активация ферментов снижается, частота циклов уменьшается, а миозиновыеые головки остаются прикрепленнымиыми к актиновымым нитям в течение всё большей части цикла. . За счёт этого напряжение мышцыы сохраняется (или «защёлкивается»). .
•От нервныхых волокон или гормональныхых источников требуются лишь слабыеые поддерживающие сигналыы.
Симпатические и парасимпатические влияния, в основном, противоположныеые:
Если одни нервыы стимулируютмулируют, , тоо другие —– тормозятмозят функциии внутренних органов.

МЕХАНИЗМАНИЗМ ДЕЙСТВИЯЙСТВИЯ МЕДИАТОРОВИАТОРОВ НАНА ГЛАДКИЕДКИЕ МЫШЦЫШЦЫ
СИМПАТИЧЕСКИЙ ОТДЕЛ
(ННОРАДРЕНАЛИНДРЕНАЛИН))
ПАРАСИМПАТИЧЕСКИЙ
ОТДЕЛ (ААЦЕТИЛХОЛИНТИЛХОЛИН))
АдренорецепторыАдренорецепторы
(альфафа ии |
) |
бетаета) |
ММ-холинорецепторынорецепторы
(ММ. |
1 |
ММ> |
2 |
ММ.)) |
|
|
3 |
Все постсинаптические рецепторыы гладких мышц
к медиаторам вегетативных нервов —–
МЕТАБОТРОПНЫЕОПНЫЕ
Действуют через внутриклеточнуюную
системустему вторыхых посредниковредников. .
Вызывают изменения проницаемости мембран, метаболизмааболизма,
синтеза веществ, ритма клеточного деления


НЕРВНАЯ СИСТЕМА
I. ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА:
СПИННОЙ МОЗГ, ГОЛОВНОЙ МОЗГ
II. ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА:
КОРЕШКИ, ГАНГЛИИ, СПЛЕТЕНИЯ, НЕРВНЫЕ СТВОЛЫ, ПЕРИФЕРИЧЕСКИЕ НЕРВЫ

ТРИ ОСНОВНЫХ ОТДЕЛА ЦНС
•СЕНСОРНАЯ СИСТЕМА
•СОМАТИЧЕСКАЯ (ДВИГАТЕЛЬНАЯ) СИСТЕМА
•ВЕГЕТАТИВНАЯ СИСТЕМА:
–СИМПАТИЧЕСКАЯ СИСТЕМА
–ПАРАСИМПАТИЧЕСКАЯ СИСТЕМА

ОСНОВНЫЕ ФУНКЦИИ ЦНС
•Регуляция двигательных функций
•Регуляция функций внутренних органов
•Восприятие, переработка и хранение информации
•Осуществление всех видов психической деятельности человека

ОСНОВНОЙ ПРИНЦИП ДЕЯТЕЛЬНОСТИ ЦНС РЕФЛЕКТОРНЫЙ
РЕФЛЕКС – ответная реакция
организма на раздражители внешней и внутренней среды с участием нервной системы.

РЕФЛЕКТОРНАЯ ДУГА - совокупность структур, необходимых для осуществления рефлекса
РЕФЛЕКТОРНАЯ ДУГА СОСТОИТ ИЗ ПЯТИ ЧАСТЕЙ:
1.Сенсорный рецептор
2.Чувствительный (афферентный) путь
3.ЦНС (нервные центры спинного и головного мозга)
4.Двигательный (эфферентный) путь
5.Рабочий орган

ФИЗИОЛОГИЯ
НЕЙРОНА
Вставочные
нейроны
Афферентные (чувствительные) нейроны
Эфферентные (двигательные) нейроны
50 тысяч разных типов нейронов в нервной системе
Рецепторы (сенсоры)

КЛАССИФИКАЦИЯ НЕЙРОНОВ
•Учитывая размеры и форму:
большие пирамидные, малые пирамидные и др.
•По количеству отростков:
униполярные, биполярные, мультиполярные.
•По длине аксона:
длинноаксонные, короткоаксонные, безаксонные.
•По типу медиатора:
адренергические, холинергические и многие др.
•По типу влияния: возбуждающие, тормозные.
•По функции:
афферентные, эфферентные, вставочные.
•По электрофизиологическим свойствам:
импульсные, безымпульсные; высокопороговые, низкопороговые и т.д.

DENDRITESДендриты NEURONНЕЙРОН
Тело
нSOMAейрона
|
|
|
|
|
|
|
|
Аксонный холмик |
|
|
NERVE |
|
|||
|
AXONAL HILLOCK |
|
|
|
|
|
|
|
|
|
|
|
Нервное |
||
|
|
|
|
||||
|
|
|
AXON |
|
TERMINAL |
||
|
|
|
|
окончание |
|||
|
|
|
Аксон |
|
|||
|
|
|
|
|
|
|
|
•ТЕЛО: метаболизм, синтез, восприятие сигналов
•ДЕНДРИТЫ: восприятие сигналов
•АКСОННЫЙ ХОЛМИК: генерация ПД
•АКСОН: передача импульсов (ПД)
•НЕРВНОЕ ОКОНЧАНИЕ: выделение медиатора
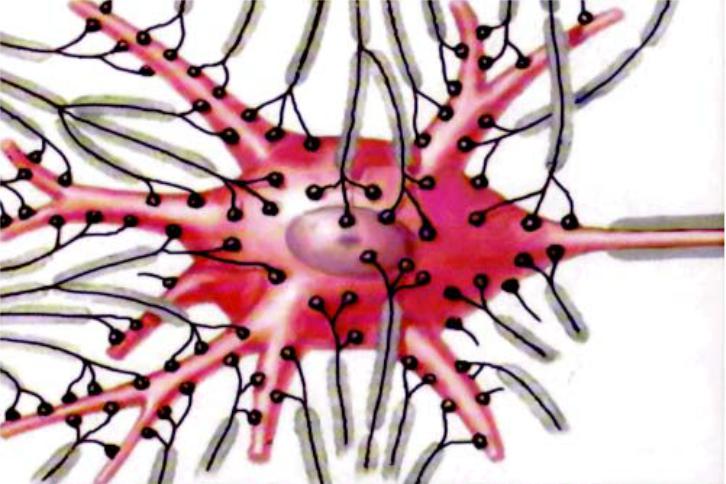
Нейроны связаны между собой многочисленными химическими синапсами (от 200 до 200 000 синапсов).
95% синапсов расположено на дендритах, и только 5% синапсов – на теле нейрона.

КЛАССИФИКАЦИЯ СИНАПСОВ
•По локализации:
аксо-сомальные, аксо-дендритные, аксоаксональные и др.
•По типу влияния: возбуждающие и тормозные
•По типу медиатора:
адренергические, серотонинергические, дофамин-ергические и многие др.
•По механизму передачи сигнала: электрические, химические, электрохимические.

ЭЛЕКТРИЧЕСКИЙ |
ХИМИЧЕСКИЙ |
СИНАПС |
СИНАПС |
Ионные |
|
токи |
|
|
ПД |
МЕДИАТОР
Ионные каналы ИОНЫ

СРАВНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА ЭЛЕКТРИЧЕСКИХ И ХИМИЧЕСКИХ СИНАПСОВ
ЭЛЕКТРИЧЕСКИЙ СИНАПС |
|
|
ХИМИЧЕСКИЙ СИНАПС |
|||
|
|
|||||
|
|
|
|
|
|
|
Без задержки |
|
|
Синаптическая задержка |
|||
Без утомления |
|
|
Быстро утомляются |
|||
Слабо чувствительны к |
|
|
Очень чувствительны к |
|||
гипоксии |
|
|
|
гипоксии |
||
Без трансформации ритма |
|
|
С обязательной трансфор- |
|||
возбуждения |
|
|
|
мацией ритма |
||
2-сторонняя передача |
|
|
1-сторонняя передача |
|||
Только возбуждающие |
|
|
Возбуждающие и тормозные |
|||
Без последействия |
|
|
Обязятельно последействие |
|||
(не сохраняют следов |
|
|
|
(сохраняют следы |
||
предшествующих |
|
|
|
предшествующих |
||
воздействий) |
|
|
|
воздействий) |
||
|
|
|
Обработка информации, |
|||
|
|
|
координация нервной дея- |
|||
|
|
|
тельности, |
память !!! |
||
|
|
|
|
|
|
|

МЕХАНИЗМ ВОЗБУЖДЕНИЯ НЕЙРОНА
ВОЗБУЖДАЮЩИЙ ПОСТСИНАПТИЧЕСКИЙ ПОТЕНЦИАЛ (ВПСП)
EPSPВПСП
10 мВmV
10 msecмсек
Свойства ВПСП:
1.Зависит от количества медиатора
2.Способен к суммации
3.Распространяется с затуханием
4.Увеличивает возбудимость нейрона, деполяризует аксонный холмик
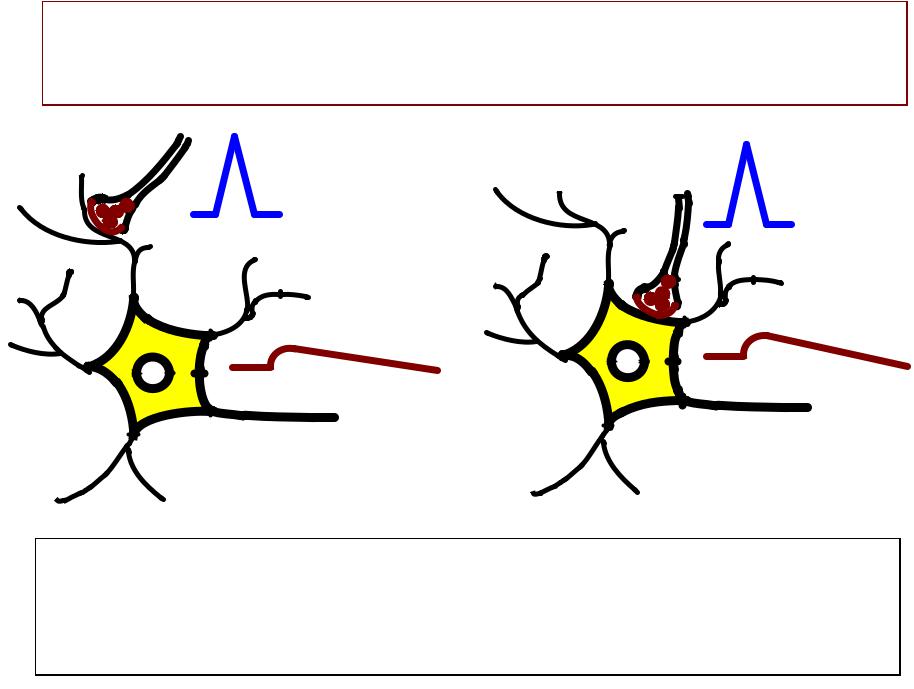
РЕАКЦИЯ НЕЙРОНА НА ОДИНОЧНЫЙ НЕРВНЫЙ ИМПУЛЬС
miniature |
miniature |
Миниатюрный |
Миниатюрный |
постсинаптиче- |
постсинаптиче- |
скийEPSPпотенциал |
скийEPSPпотенциал |
Слишком слабая деполяризация постсинаптической мембраны (0.1-1 мВ).
ПД не возникает.

РЕАКЦИЯ НЕЙРОНА НА СЕРИЮ НЕРВНЫХ ИМПУЛЬСОВ
серия ПД
ВПСП
EPSP10 мВ – результат
временной суммации
subthreshold
ВПСП распространяется с затуханием, вызывает допороговую деполяризацию аксонного холмика. ПД не возникает.
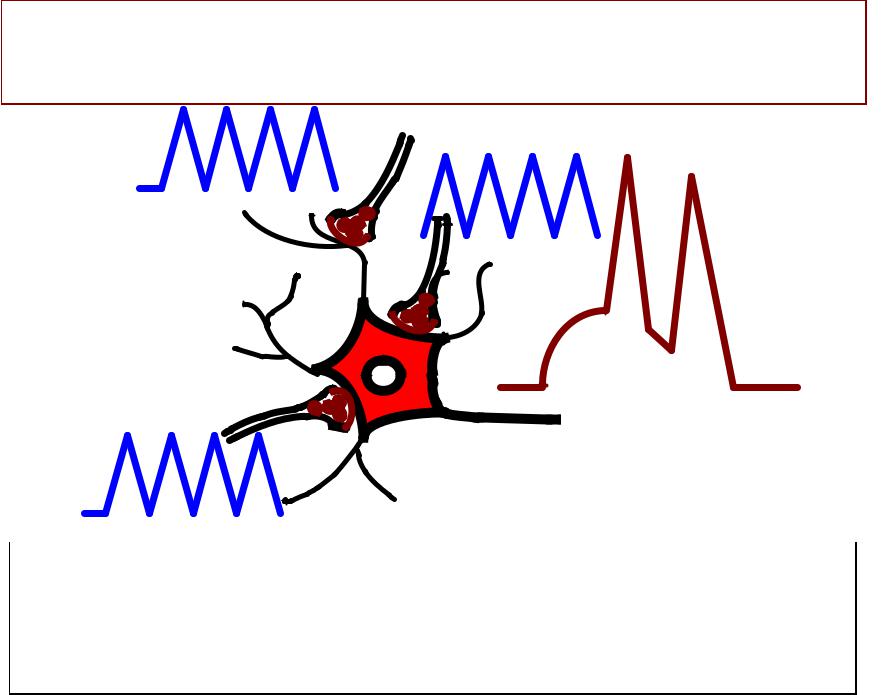
РЕАКЦИЯ НЕЙРОНА НА ВПСП, ВОЗНИКШИЕ ОДНОВРЕМЕННО В РАЗНЫХ СИНАПСАХ
Пороговая деполяризация
аксонногоEPSP холмика –
результат пространствен-
threshold
ной суммации
Множество ВПСП, возникших одновременно в
разных участках нейрона, вызывают пороговую деполяризацию аксонного холмика.
Происходит генерация ПД.
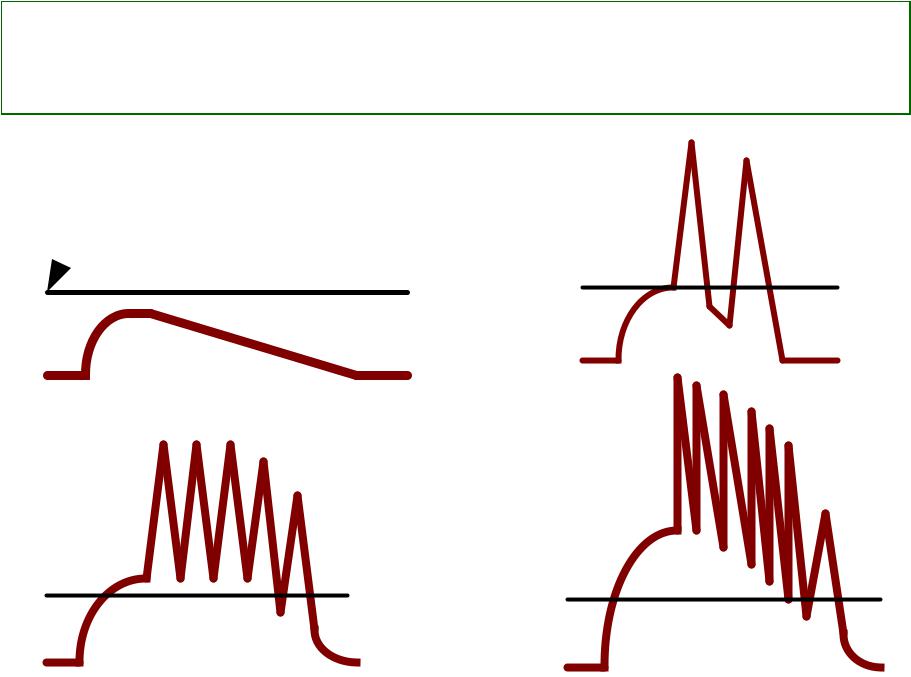
Чем выше деполяризация аксонного холмика, тем больше частота импульсов
Критическийcritical levelуровень
 ofдеполяризацииdepolarization
ofдеполяризацииdepolarization

СВОЙСТВА ТПСП:
1.Зависит от количества медиатора
2.Способен к суммации
аксон 3. Распространяется с затуханием
4.Уменьшает возбудимость нейрона, вызывает гиперполяризацию аксонного холмика
Тормозной медиатор - ГЛИЦИН

ПОСТСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
•Сущность постсинаптического торможения – гиперполяризация нервной клетки.
•Гиперполяризация возникает за счёт входа в клетку ионов хлора и/или выхода из клетки ионов калия.
•Гиперполяризация приводит к увеличению порогового потенциала (дельта-V) в области аксонного холмика. Возбудимость нейрона при этом снижается.
•Заторможенный нейрон перестаёт реагировать на любые поступающие к нему импульсы.

ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
ПД
ТПСП
Аксон тормозного нейрона
ПД
EPSPВПСП
Торможение развивается в пресинаптическом нервном
окончании. ПД не может пройти к нейрону через заблокированный участок одного из многих пресинаптических входов.
Тормозной медиатор – ГАМК (гамма-аминомасляная кислота)

ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
•Пресинаптическое торможение развивается за счёт длительной стойкой деполяризации постсинаптической мембраны в аксо-аксональном синапсе.
•Длительная деполяризация приводит к инактивации натриевых каналов и блокаде проведения импульсов к нейрону по возбуждающему нервному волокну.
•Возбудимость нейрона при этом не меняется. Нейрон продолжает реагировать на импульсы, поступающие к нему по другим нервным волокнам.

ВЫВОД
ТОРМОЖЕНИЕ – это активный нервный процесс, который направлен на прекращение генерации импульсов и (или) выделения медиатора из нервных окончаний.
Торможение всегда является следствием возбуждения.
НЕЙРОМЕДИАТОРЫ

КЛАССИФИКАЦИЯ МЕДИАТОРОВ
•Низкомолекулярные, кратковременного действия:
–АЦЕТИЛХОЛИН
–АМИНЫ
–АМИНОКИСЛОТЫ
–ОКСИД АЗОТА (NO)
•Высокомолекулрные, долговременного действия:
–НЕЙРОПЕПТИДЫ
•Гипоталамические
•Гипофизарные
•Гастроинтестинальные
•и другие
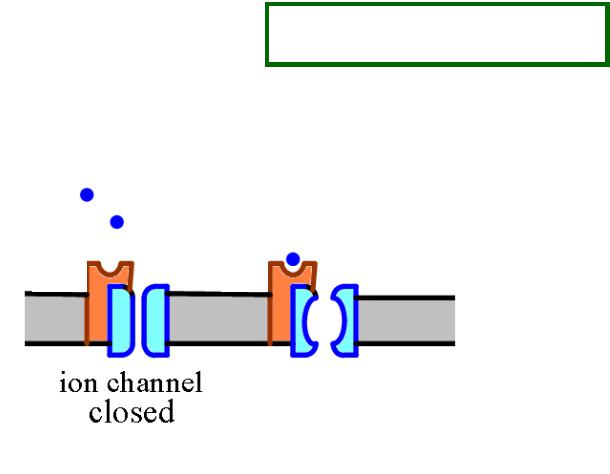
МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
ИОНОТРОПНЫЕ
(связанные с хемочувствительными ионными каналами)
Р Р
ИОННЫЙ |
ИОННЫЙ |
КАНАЛ |
КАНАЛ |
ЗАКРЫТ |
ОТКРЫТ |
|
|
С ионотропными рецепторами заимодействуют:
ацетилхолин, глутамат, аспартат, глицин, ГАМК

МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
МЕТАБОТРОПНЫЕ
(связанные с системой вторых посредников)
С метаботропными рецепторами взаимодействуют:
катехоламины, серотонин, ГАМК, опиоиды,
нейропептиды и др.

МОЛЕКУЛЯРНЫЕ РЕЦЕПТОРЫ ПОСТСИНАПТИЧЕСКИХ МЕМБРАН
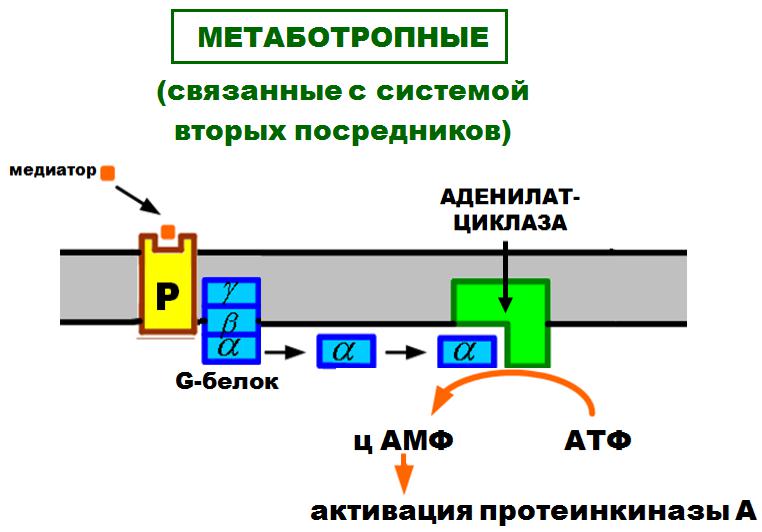
МЕТАБОТРОПНЫЕ
(связанные с системой вторых посредников)
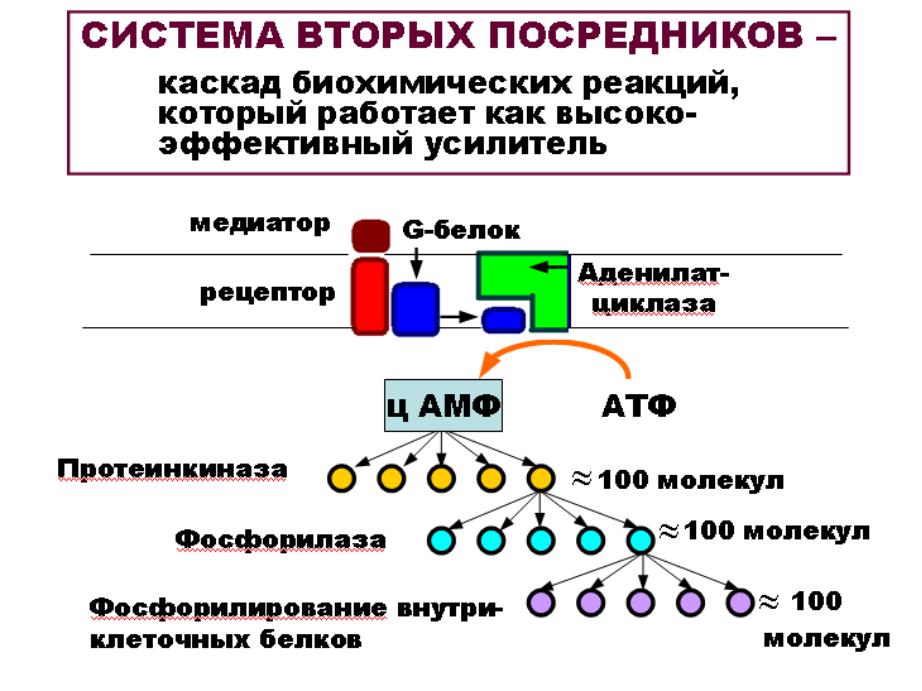
АДЕНИЛАТЦИКЛАЗА
Р
G-белок
ц АМФ |
АТФ |
||
|
|
|
|
|
|
|
|
активация протеинкиназы А

АКТИВАЦИЯ СИСТЕМЫ ВТОРЫХ ПОСРЕДНИКОВ
•Медиатор (первый посредник) химически связывается
срецептором постсинаптической мембраны.
•Происходит активация G-белка.
•Свободная альфа-субъединица G-белка активирует фиксированный в мембране фермент – аденилатциклазу.
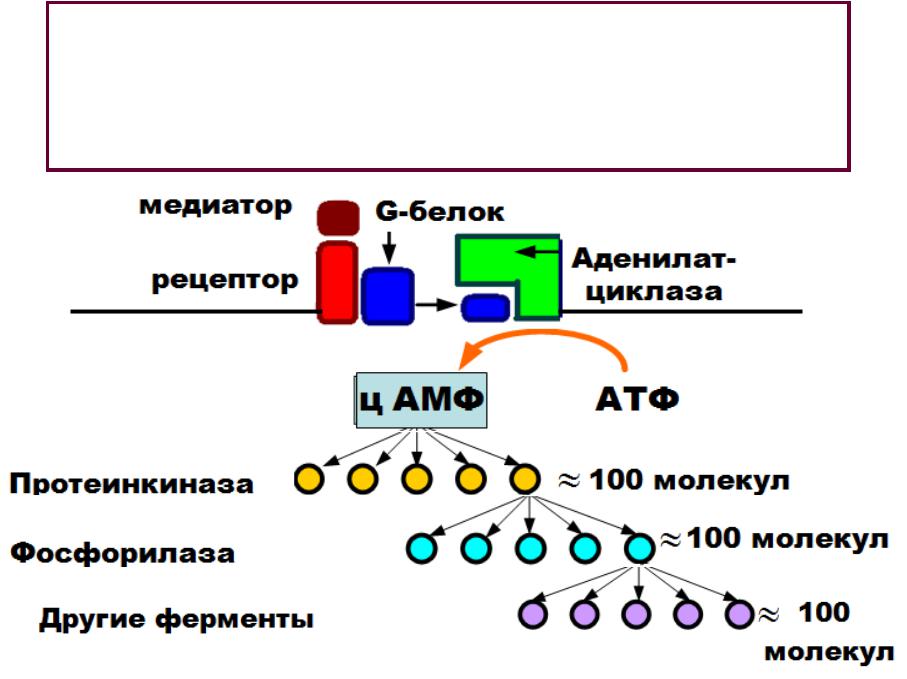
•Фермент необходим для образования ц АМФ.
•ц АМФ и является внутриклеточным (вторым) посредником, который активирует внутриклеточный фермент - фосфорилазу.
•Фосфорилирование разных белков вызывает изменение свойств клетки:
–открытие ионных каналов;
–изменение иетаболизма;
–изменение ритма клеточного деления и др.