

Матричные биосинтезы №1 Репликация. Репарация. Транскрипция
.pdf21
1. Инициация - узнавание
и прочное |
связывание |
||
фермента, |
образование |
||
первой |
фосфодиэфирной |
||
связи |
между |
пурин- |
|
трифосфатом |
(АТФ или |
||
ГТФ) |
и |
следующим |
|
нуклеотидом. |
|
|
|
2. |
Элонгация |
- |
|
последовательное наращивание цепи РНК.
Скорость элонгации 40-50 нукл./сек.
3. Терминация - в терминаторе присутствует палиндром. В синтезируемой РНК формируется шпилька. Шпилька меняет конформацию РНК-полимеразы и фермент теряет сродство к ДНК.
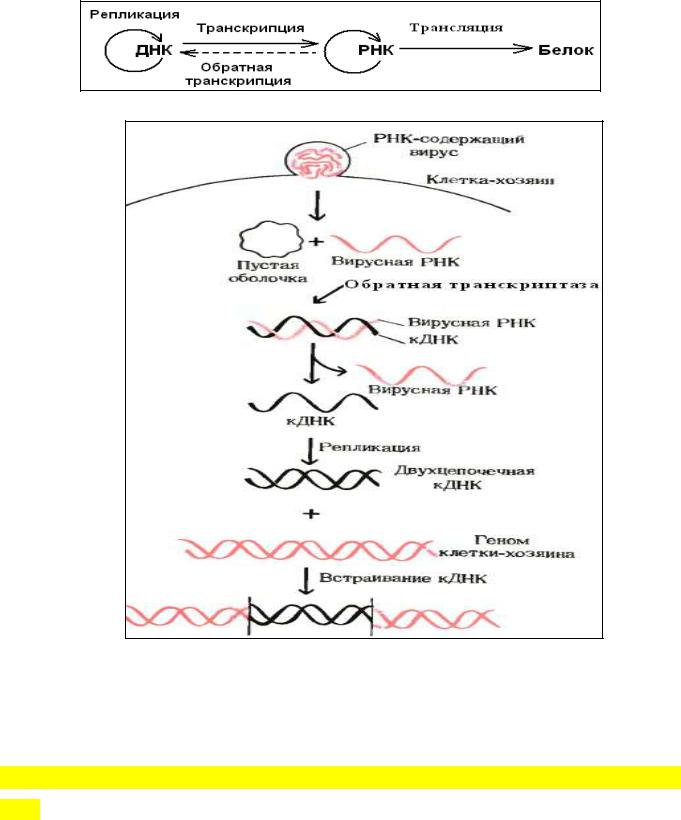
Обратная транскрипция
Некоторые онкогенные РНК-содержащие вирусы животных имеют уникальный фермент - РНК-зависимую ДНК-полимеразу - обратную транскриптазу и способны встраивать генетическую информацию со своей вирусной РНК в ДНК генома клетки-
хозяина.
После того как вирус попадает в клетку-хозяина, этот фермент способен катализировать синтез ДНК, комплементарной по отношению к вирусной РНК,
которая играет при этом роль матрицы. В результате образуется кДНК, которая может в течение многих поколений оставаться в скрытом, т.е. неэкспрессируемом,
состоянии. При определенных условиях такие бездействующие вирусные гены могут активироваться и вызывать репликацию вируса; при других же условиях они могут способствовать превращению клеток в раковые. Открытие обратной транскриптазы позволило по-новому сформулировать центральную догму молекулярной биологии.
22
Рис. Участие обратной транскриптазы в образовании к-ДНК на вирусной
одноцепочечной РНК-матрице в животной клетке
4. Процессинг РНК: посттранскрипционные превращения различных типов РНК
Синтезированные молекулы РНК являются функционально неактивными и в
дальнейшем претерпевают ряд ферментативных превращений, которые называют
23
посттранскрипционным процессингом. У эукариот процессингу подвергаются все виды пре-РНК, у прокариот – только предшественники рРНК и тРНК.
Процессинг предшественника рРНК. Предшественники рРНК являются более крупными молекулами по сравнению со зрелыми рРНК.
Упрокариот большая прерибосомная 30S-РНК расщепляется специфичными нуклеазами с образованием 5S-рРНК, 16S-рРНК, и 23S-рРНК.
Уэукариот большая прерибосомная 45S-РНК расщепляется специфичными нуклеазами с образованием 5,8S-рРНК, 18S-рРНК, и 28S-рРНК.
рРНК как эукариотических, так и прокариотических клеток образуются из более длинных молекул – предшественников, называемых прерибосомными РНК. У
прокариот 16S- и 23S-р-РНК образуются из одного длинного 30S-предшественника,
молекулярная масса которого составляет приблизительно 2×106. Этот предшественник метилируется по специфическим основаниям и расщепляется, давая
17S- и 25S-промежуточные РНК, которые затем процессируются путем отщепления остатков с помощью нуклеаз, образуя характерные для прокариот 16S- и 23S-р-РНК.
5S-р-РНК образуется отдельно из 3-концевого участка 30S-предшественника.
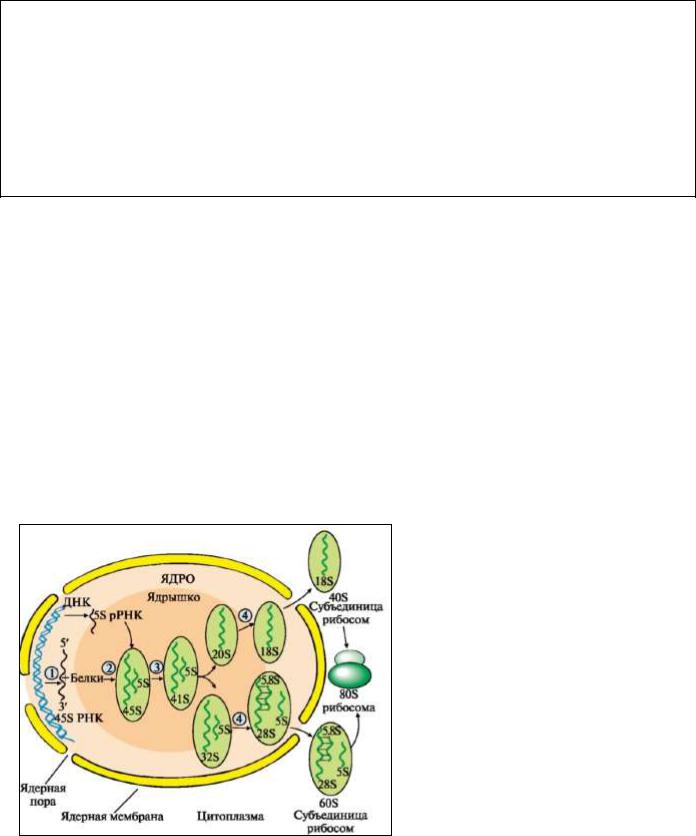
Рис. Посттранскрипционные модификации прерРНК и образование рибосом
1 - транскрипция пре-рРНК;
2 - связывание 45SрРНК с белками и 5S-рРНК;
3 - метилирование пре-рРНК и расщепление на отдельные фрагменты;
4 - дальнейшее укорочение рРНК и
формирование 40S- и 60S-субъединиц рибосом

24
У эукариот 18S- и 28S-р-РНК образуются в несколько этапов: из большой 45S-
прерибосомной РНК. Процессинг протекает в ядрышке. Сначала происходит метилирование более чем 100 нуклеотидов 45S-предшественника. Затем метилированная 45S-РНК претерпевает ряд ферментативных расщеплений,
приводящих в конечном итоге к появлению 18S-, 28S- и 5,8S-р-РНК, характерных для эукариотических рибосом. 5S-р-РНК синтезируется отдельно.
Процессинг предшественника тРНК. 1. Формирование на 3’-конце последовательности Ц-Ц-А. Для этого у одних пре-тРНК с 3’-конца удаляются лишние нуклеотиды до «обнажения» триплета Ц-Ц-А, у других идет присоединение
этой последовательности.
2.Формирование антикодоновой петли происходит путем сплайсинга и удаления интрона в средней части пре-тРНК.
3.Модификация нуклеотидов путем дезаминирования, метилирования,
восстановления. Например, образование псевдоуридина и дигидроуридина.
тРНК также образуются из более длинных РНК-предшественников в результате ферментативного удаления лишних нуклеотидов с 5- и 3-концов молекулы. В ходе процессинга в предшественниках т-РНК происходят изменения двоякого рода. Во-первых, к некоторым т-РНК присоединяется 3-концевая тринуклеотидная последовательность -С-С-А (3-); в других т-РНК этот 3-концевой тринуклеотид уже содержится в транскрипте. 3-концевой остаток А представляет собой именно ту часть молекулы т-РНК, с которой ковалентно связывается соответствующая ей аминокислота перед включением в растущую полипептидную цепь на рибосоме. Во-вторых, ряд оснований в т-РНК специфическим образом модифицируется: одни метилируются, другие дезаминируются, третьи восстанавливаются. Модифицированные основания располагаются во всех т-РНК в определенных положениях.
25
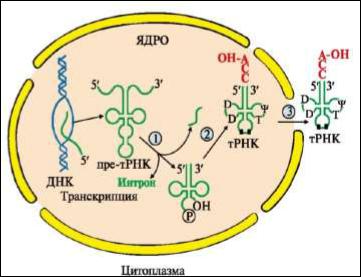
Рис. |
Процессинг транскриптов т- |
||
РНК: |
|
|
|
1 |
- |
удаляются |
участки |
полинуклеотидной цепи на 5'- и 3'-
концах молекулы пре-тРНК и интрон в центральной области молекулы;
2 - модифицируются азотистые основания, к 3-концу присоединяется
триплет ССА; 3 - в цитоплазму
выходят зрелые тРНК
Особенности транскрипции у эукариот
У эукариот процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция - в цитоплазме). У эукариот существуют специализированные РНК-полимеразы. В ядре выделяют 3 типа РНК-
полимераз:
РНК-полимераза I - синтезирует rРНК (кроме 5S rРНК).
РНК-полимераза II - синтезирует mРНК и некоторые sРНК.
РНК-полимераза III - синтезирует tРНК, некоторые sРНК и 5SrРНК.
РНК-полимеразы различаются количеством субъединиц, их аминокислотным составом, и зависимостью от катионов магния и марганца. Помимо ядерных РНК-
полимераз у эукариот есть еще РНК-полимеразы хлоропластов и митохондрий. Они кодируются в ядре, а не в соответствующих органеллах. В органеллах образуются свои тРНК, рРНК и рибосомные белки.
Процессинг (созревание) рРНК и тРНК у эукариот принципиально не отличается от такового у прокариот. Процессинг мРНК сильно отличается и состоит из нескольких этапов.
1.Кепирование 100% мРНК
2.Полиаденилирование ~95% мРНК

26
3.Сплайсинг ~95% мРНК. Сплайсингу подвергаются только полиаденилированные mРНК.
4.Редактирование. Показано лишь для нескольких мРНК.
Процессинг предшественника мРНК 1. Кэпирование (англ. cap - шапка) –
происходит еще во время транскрипции, состоит в том, что к 5’-трифосфату концевого нуклеотида пре-мРНК присоединяется 5’-углерод N 7-метил-гуанозина. «Кэп» необходим для защиты молекулы РНК от 5’-3’-экзонуклеаз.
2. При транскрипции зон ДНК, несущих информацию о белках, образуются гетерогенные ядерные РНК, по размеру намного превосходящие мРНК. Дело в том,
что из-за мозаичной структуры генов эти гетерогенные РНК включают в себя информативные (экзоны) и неинформативные (интроны) участки. При особом процессе – сплайсинге (англ. splice – склеивать встык) происходит удаление интронов
исохранение экзонов.
3.Полиаденилирование – при помощи полиаденилат-полимеразы с использованием молекул АТФ происходит присоединение к 3’-концу 100-200
адениловых нуклеотидов, формирующих поли (А)-хвост.
Все стадии процессинга мРНК происходят в РНП-частицах
(рибонуклеопротеидных комплексах). По мере синтеза про-мРНК, она тут же образует комплексы с ядерными белками - информоферами. И в ядерные, и в цитоплазматические комплексы мРНК с белками (информосомы) придают РНК необходимую конформацию и защищают. В составе информосом mРНК может жить от нескольких минут до нескольких дней, не подвергаясь действию нуклеаз (мРНК живут неделями в ооцитах, предшественниках яйцеклеток).
Полисома - комплекс мРНК с несколькими или многими рибосомами.

27
Кэпирование -
надевание "шапочки".
"Сар" представляет собой метилированный ГТФ,
присоединенный в необычной позиции 5'-5' и
две метилированные
рибозы в первых двух нуклеотидах mРНК. По мере образования про-mРНК (еще до
30-ого нуклеотида), к 5'-концу, несущему пуринтрифосфат, присоединяется гуанин,
после чего происходит метилирование.
ГТФ + гуанинтрансфераза (Е)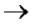 Е~фГ + ф-ф
Е~фГ + ф-ф
5'ф-ф-ф-Пур-ф-Х-ф-Y-ф-Z-...+E~фГ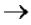 Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...
Г5'-ф-ф-ф-5'-Пур-ф-Х-ф-Y-ф-Z...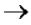
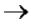 (метилирование) Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
(метилирование) Гm-ф-ф-ф-Пурm-ф-Хm-ф-Y-ф-Z-...
Назначение "Сар"
1.Защита 5'-конца мРНК от действия экзонуклеаз.
2.За счет узнавания "Сар"-связывающими белками происходит правильная установка мРНК на рибосоме.
Полиаденилирование. Когда синтез про-мРНК завершен, то на расстоянии примерно 20 нуклеотидов в направлении к 3' - концу от последовательности 5'-
AAУAA-3' происходит разрезание специфической
эндонуклеазой и к новому 3'-концу присоединяется от 30 до 300 остатков АМФ
(безматричный синтез). Каждый вид мРНК имеет "поли-А хвост" определенной длины. Он защищает 3'-конец от гидролиза, т.к. покрыт полиА-связывающими белками. В значительной степени время жизни мРНК коррелирует с длиной полиА-
28
хвоста. мРНК ряда генов не полиаденилируется (например, гистоновых генов).
Полиаденилированные про-мРНК подвергаются сплайсингу.
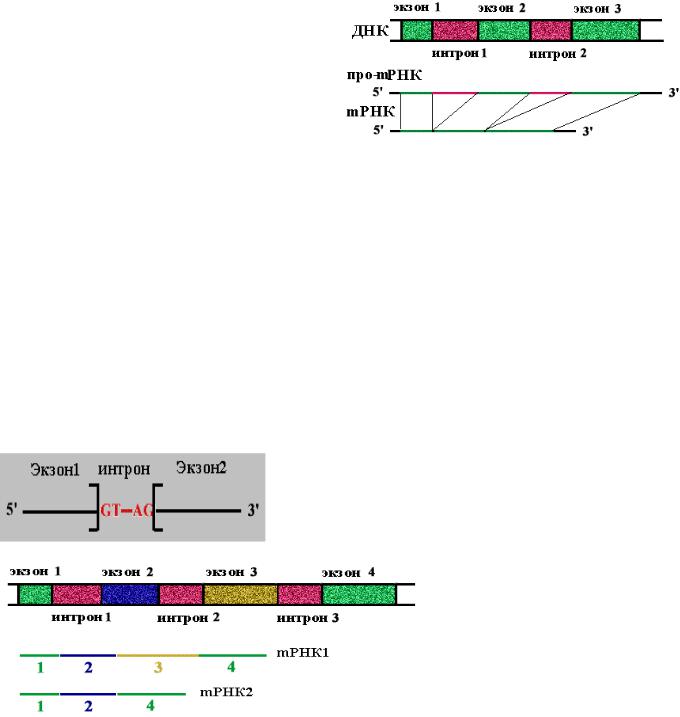
Сплайсинг - вырезание копий интронов (некодирующих участков гена) из про-
мРНК и сшивание копий экзонов (кодирующих участков гена) с образованием зрелой мРНК.
Явление сплайсинга РНК (от англ. to splace - сшивать без узлов) открыл в 1978
г. Филипп Шарп (Массачусетский технологический институт).
На долю интронов приходится в 5-7
раз больше нуклеотидных пар, чем на долю экзонов. Количество экзонов в гене больше, чем интронов.
Копии интронов гидролизуются до нуклеотидов. Сплайсинг показан для большинства мРНК и некоторых тРНК. У простейших найден автосплайсинг рРНК.
Сплайсинг показан даже для архебактерий.
Существуют разные механизмы сплайсинга, его осуществляют ферменты-
матюразы. В некоторых случаях в процессе сплайсинга участвуют sРНК. В случае автосплайсинга процесс происходит благодаря третичной структуре про-РНК.
В м-РНК 5' и 3' концы интрона очень консервативны: 5'(ГT-интрон-AГ)3'. При сшивании копий экзонов соблюдается порядок их расположения в гене.
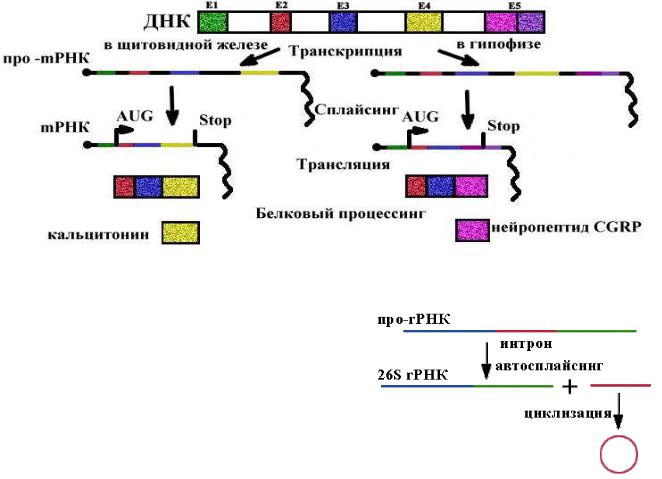
29
Альтернативный сплайсинг мРНК кальцитонинового гена у млекопитающих
Во всех клетках есть кальцитониновый ген, но в клетках щитовидной железы он экспрессируется в виде гормона кальцитонина, а в клетках гипофиза -
нейропептида CGRP (пептида, имеющего отношение к гену кальцитонина). Ген один,
а белки получаются разные в результате сплайсинга мРНК и процессинга полипептидов. В клетках других тканей этот ген не экспрессируется.
Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в
которых помимо ферментов, вырезающих и сшивающих участки про-мРНК, имеются белки, придающие про-мРНК нужную конформацию.
|
Автосплайсинг открыт Томасом Чеком |
(США) в 1982 |
|||
году. |
Впервые |
было |
показано, |
что |
|
каталитической |
активностью обладают |
не |
|
||
только белки. |
|
|
|
|
|
|
РНК-зимы - РНК с каталитической активностью. |
|
|||
Малые РНК (мяРНК) обнаружены в количестве 103-105 копий на клетку. Поскольку в большинстве случаев эти РНК обогащены урацилом, они
называются U1, U2... Их размер от 100 до 300 нукл. Все они кодируются в ядре, но работают как в ядре (small nuclear - SN), так и в цитоплазме (small cytoplasmic - SC).
Рибонуклеопротеидные комплексы в ядре участвуют в полиаденилированиии и сплайсинге, РНП в цитоплазме входят в состав информосом. Малая РНК U4
30
присутствует в комплексах, участвующих в полиаденилировании. Если получить антитела к белкам, связывающимся с U4, то не происходит полиаденилирования и сплайсинга. При красной волчанке (аутоимунном заболевании) вырабатываются антитела к белкам комплекса с U4.
5. Генетический код и его свойства. Значение тРНК в декодировании генетической информации
Ген - это участок ДНК, кодирующий одну полипептидную цепь или одну молекулу тРНК, рРНК или мяРНК. Гены тРНК, рРНК, мяРНК белки не кодируют.
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности нуклеотидов в
ДНК. Поскольку ДНК непосредственного участия в синтезе белка не принимает, то код записывается на языке РНК. В РНК вместо тимина входит урацил.
Свойства генетического кода
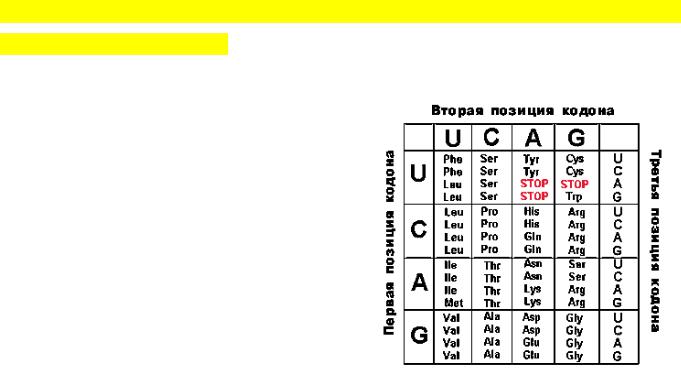
1.Триплетность. Каждая аминокислота кодируется последовательностью из 3-х
нуклеотидов.
2.Вырожденность. Все аминокислоты, за исключением метионина и триптофана,
кодируются более чем одним триплетом. Всего 61 триплет кодирует 20 аминокислот.
3. Однозначность. Каждый триплет кодирует лишь одну аминокислоту или является терминатором трансляции.
4. Наличие межгенных знаков препинания. В конце каждого гена, кодирующего полипептид, находится, по меньшей мере, один из 3-х терминирующих кодонов, или стоп-сигналов: UAA, UAG, UGA. Они терминируют трансляцию. Условно к знакам