

Матричные биосинтезы №1 Репликация. Репарация. Транскрипция
.pdf11
Оrigin (ori) - район начала репликации ДНК, содержит
4 шпильки, которые опознаются РНК-полимеразой и используются в качестве матрицы. По мере образования РНК шпилька плавится. Образуется РНК-затравка длиной 24
нуклеотида, 3'-конец которой используется ДНК полимеразой III.
Реплисома – комплекс ферментов для синтеза ДНК
(ДНК-полимераза III, ДНК-полимераза I, лигаза).
Репликон - участок ДНК между двумя участками ori. Каждая эукариотическая хромосома – полирепликон.
По мере расплетания и движения репликативной вилки на нити открываются участки, где возможен синтез новой нити в направлении 5' → 3'.
Направление 5' → 3' другой материнской нити ДНК совпадает с направлением движения репликативной вилки. Поэтому синтез дочерней нити (в направлении 5' → 3') возможен только после расплетания части ДНК и освобождения участка для синтеза. Таким образом, синтез дочерней ДНК на одной из нитей материнской ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
В целом для синтеза ДНК необходим ряд ферментов.
12
Топоизомеразы - ферменты, изменяющие топологию ДНК. Топоизомеразы меняют число зацеплений одной цепи за другую. Делятся на два класса: тип I (релаксазы) - уменьшают число зацеплений, тип II
(гиразы) - увеличивают число зацеплений.
Гиразы вносят двуцепочечный разрыв ДНК по принципу работы рестриктаз.
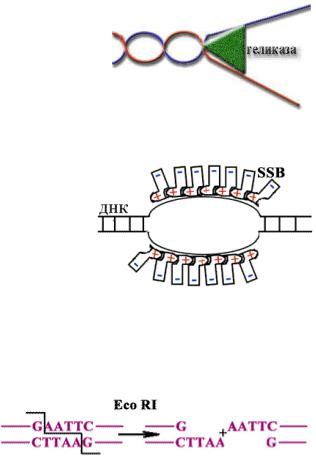
Хеликазы - ферменты, денатурирующие (расплетающие) ДНК.
SSB белки (single strand bind) - связываются с одноцепочечной ДНК электростатически, когда в ДНК образуется расплавленный участок, не дают цепям схлопнуться. Они не денатурируют ДНК, а лишь фиксируют, удерживают матричные цепи ДНК в репликативной вилке.
Праймаза – фермент, создающий РНК-затравку для начала синтеза ДНК. У
эукариот РНК-затравки размером 6-10
нуклеотидов удаляются РНК-азой. Бреши заделываются репарирующими ферментами.
Рестриктазы - эндонуклеазы, узнающие определенные последовательности,
делают разрезы в обеих цепях ДНК, затем поворачивают концы ДНК на 360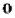 и
и
проявляет лигазную активность, т.е. сшивают цепи ДНК. При этом используется энергия АТФ. Результат деятельности гираз - супервитки. Суперспирализованная ДНК напряжена. В некоторых местах цепи расходятся, т.к. расплавляются А-Т
участки.
Ферменты репликации эукариот и их функция
13
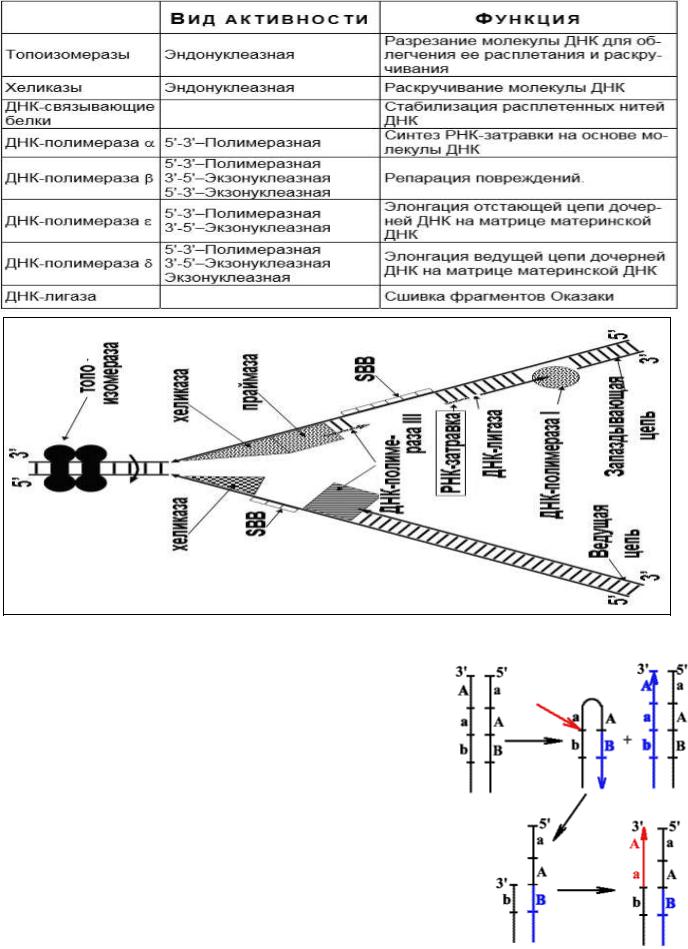
Проблема репликации концов линейных молекул ДНК эукариот
Удаление РНК-праймеров после завершения синтеза линейных ДНК в виде фрагментов Оказаки и заделывание образующихся между фрагментами брешей нуклеотидами ДНК приводит к тому, что дочерние цепи ДНК оказываются короче материнских на размер первого РНК-праймера (10-20 нукл.).
Образуются 3'-оверхенги, т.е. выступающие 3'-концы материнских цепей. Они узнаются теломеразой -
ферментом, содержащим помимо белковой части еще и РНК, выполняющую роль матрицы для наращивания
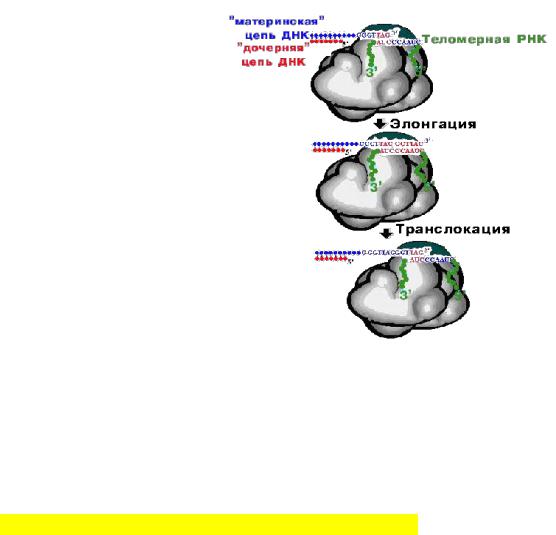
14
ДНК повторами. Теломераза последовательно наращивает материнские цепи ДНК повторами, используя 3'-оверхенги в качестве затравок. Образующиеся длинные одноцепочечные концы в свою очередь служат матрицами для синтеза дочерних цепей традиционным репликативным механизмом.
Хромосомы соматических клеток человека фланкированы многократно повторенными гексамерами TTAГГГ, общая длина районов с повторами может достигать 10 тыс. пар нуклеотидов. В
комплексе со специфическими белками такие тандемные повторы образуют
теломеры, защищающие концы ДНК от действия экзонуклеаз, предотвращающие неправильную рекомбинацию и позволяющие концам хромосом прикрепляться к ядерной оболочке.
При каждом раунде репликации происходит укорочение теломер в среднем на 50 пар нуклеотидов. Поскольку
теломерные последовательности не являются кодирующими, они выступают в роли буферной зоны - как защита от "проблемы концевой репликации". Укорочение ДНК в ходе каждого раунда репликации лишь сокращает нетранскрибируемый текст теломеры, но не приводит к утрате смысловых последовательностей - генов и регуляторов их экспрессии.
2. Основные повреждения в ДНК и их репарация
На геном любой неделящейся клетки постоянно оказывает влияние окружающая среда, при этом вполне вероятны повреждения в составе генома:
изменение нуклеотида (например, дезаминирование), сшивки азотистых оснований друг с другом, разрывы цепей, отрыв пуриновых нуклеотидов и т.п. Такие изменения быстро определяются специальными ферментами, пораженный участок удаляется экзонуклеазами, заполняется ДНК-полимеразой β и сшивается ДНК-лигазой. В

15
делящейся клетке мутации могут также возникать во время синтеза ДНК. Поэтому в клетках существует двойная система проверки точности репликации: одна непосредственно при ДНК-полимеразной реакции, другая – анализ уже синтезированной ДНК.
Причины ошибок при синтезе ДНК
In vitro происходит 1 ошибка на 100 тыс. нукл. для средней ДНК-полимеразы.
Добавление аналогов нуклеотидов, например, бромдезоксиуридина - аналога тимидина – это способ повышения количества ошибок как одно из средств борьбы со СПИДом и раком. Аналоги одинаково вредны для всех клеток, однако в пораженных вирусом клетках чаще проходит репликация.
1. Апуринизация. Каждая соматическая клетка теряет за сутки около 10000 пуринов и пиримидинов. Разрывается N-
гликозидная связь между пуриновым основанием и дезоксирибозой. В ДНК образуются АП-сайты. Причины апуринизации: изменение рН, ионизирующее излучение,
повышение температуры и т.д. Пиримидины тоже могут отщепляться, но скорость этого процесса ниже.
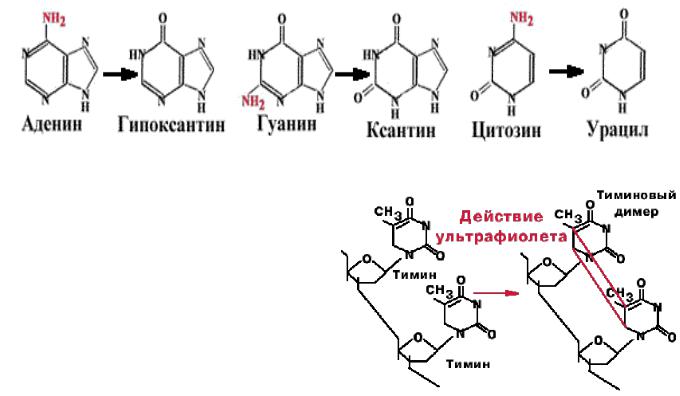
2. Дезаминирование. Аденин превращается в гипоксантин, который образует две водородные связи с цитозином. Гуанин превращается в ксантин, который образует водородные связи с тимином. При дезаминировании цитозина образуется урацил.
Тимин не может быть дезаминирован (единственный в ДНК). Наличие тимина в ДНК
(вместо урацила) позволяет отличать дезаминированнный цитозин (т.е. урацил) от законного урацила, если бы он был в ДНК. N-гликозилаза - фермент, который узнает дезаминированное основание, разрывает N-гликозидную связь и удаляет неправильное основание.
После этого АП-специфическая эндонуклеаза вносит одноцепочечный разрыв,
и фосфодиэстераза отщепляет от ДНК ту сахарофосфатную группу, к которой теперь
16
не присоединено основание. Появляется брешь размером в один нуклеотид, которую заделывает ДНК-полимераза, а лигаза сшивает концы ДНК.
ДНК - двуцепочечна в отличие от РНК. Наличие второй цепи обеспечивает исправление ошибок. Дезоксирибоза более устойчива, чем рибоза, к действию щелочи, т.е. при рН > 8, ДНК устойчива, а РНКнет.
3. Тиминовые димеры. Под действием ультрафиолетого света |
|
происходит ковалентное сшивание рядом |
|
стоящих пиримидинов. При сшивании |
|
тиминов образуется циклобутановое |
|
производное, блокирующее репликацию. |
|
Фермент фотолиаза - узнает тиминовые |
|
димеры и на свету или в темноте образует с |
ними |
комплекс. При освещении видимым светом происходит активация |
|
фермента, циклобутановое кольцо разрывается, и вновь получаются два тимина.
Этот процесс называется фотореактивацией.
И дезаминированные основания, и тиминовые димеры, кроме того, могут удаляться с помощью эксцизионной репарации. Специфические эндонуклеазы производят одноцепочечные разрезы (инцизия). Затем происходит удаление
(эксцизия) нескольких нуклеотидов и заделывание бреши. У E. сoli заделыванием бреши занимается ДНК-полимераза I. Лигаза сшивает цепь. Она же ликвидирует одноцепочечные разрывы, возникающие при действии ионизирующей радиации.
17
У E.coli эксцизионная репарация осуществляется мультиферментным комплексом,
включающим белки uvrA, uvrB, uvrC (ultraviolet repair), которые узнают поврежденный участок и вносят 5'- и 3'- разрывы с разных сторон от него, uvrD - хеликазу, которая отсоединяет вырезанный олигомер - 12 нуклеотидов, используя энергию АТФ.
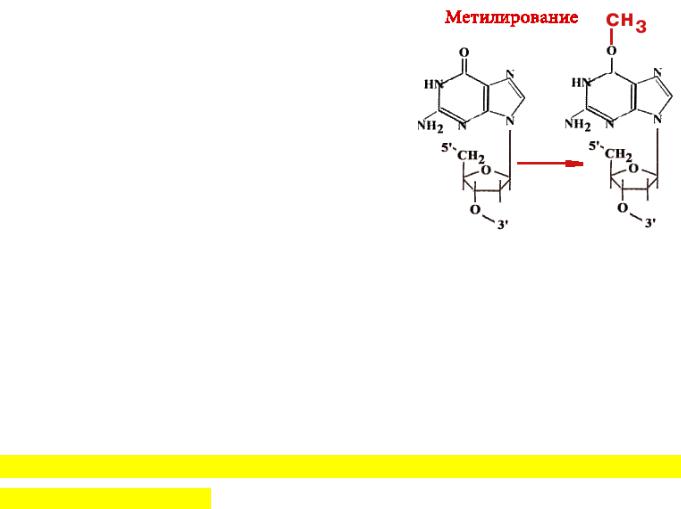
У эукариот существует функциональный (но не структурный) аналог такого мультиферментативного комплекса. Имеется 14 позиций, по которым ДНК метилируется. Гуанин может быть метилирован (по кислороду в 6-ом положении) и в такой форме будет связываться не только с цитозином, но и с тимином. Таким образом, в два шага может произойти замена пары Г-Ц на А-Т. Фермент принимает метильную группу на один из 12 цистеиновых остатков и при этом "гибнет".
3. Транскрипция – синтез РНК: субстраты, кофактор. РНК-полимераза.
Обратная транскрипция
Транскрипция (англ. transcription – переписывание) - это биосинтез всех видов РНК по матрице ДНК, осуществляемый ферментом ДНК-зависимой РНК-
полимеразой. У прокариот синтез всех видов РНК осуществляется одним и тем же ферментом. У эукариот - 3 ядерные РНК-полимеразы, митохондриальные РНК-
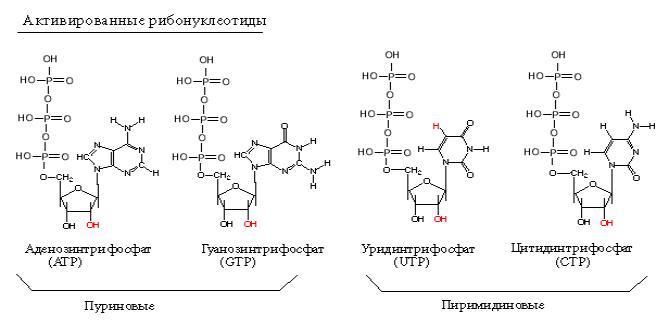
полимеразы, хлоропластные РНК-полимеразы. Субстратами для РНК-полимераз служат рибонуклеозид-трифосфаты (активированные нуклеотиды). Весь процесс транскрипции осуществляется за счет энергии макроэргических связей актвированных нуклеотидов.
18
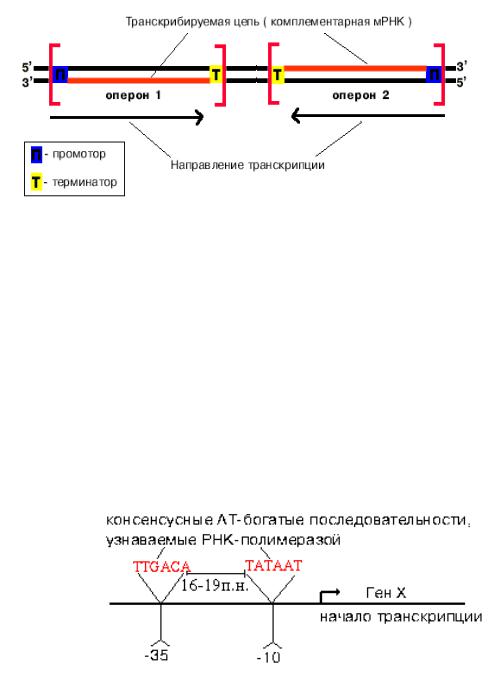
Биосинтез РНК происходит в участке ДНК, который называется транскриптом,
с одного края он ограничен промотором (начало), с другого – терминатором
(конец). Как в любом матричном биосинтезе в транскрипции выделяют 5
необходимых элементов:
•матрица – одна из цепей ДНК
•растущая цепь – РНК
•субстрат для синтеза – рибонуклеотиды (УТФ, ГТФ, ЦТФ, АТФ)
•источник энергии – УТФ, ГТФ, ЦТФ, АТФ
•ферменты – РНК-полимеразы.
Существует три основных типа РНК-полимераз: для синтеза пре-рРНК
(РНКполимераза I), для синтеза пре-мРНК (РНК-полимераза II), для синтеза пре-
тРНК и 5S-рРНК (РНК-полимераза III). В составе РНК-полимеразы E.coli выделяют четыре субъединицы: две α-субъединицы, по одной β- и β’-субъединице. Имеется также дополнительный белковый σ-фактор Последний необходим только для связывания с промотором и не участвует в удлинении цепи РНК. Строение РНК-
полимераз эукариот имеет много общего со структурой бактериального фермента:
они имеют по две больших субъединицы и несколько малых субъединиц.
Принципы транскрипции:
19
1. Комплементарность.
2.
Антипараллельность.
3.Униполярность.
4.Беззатравочность.
5.Асимметричность.
РНК синтезируется комплементарно и антипараллельно транскрибируемой цепи ДНК. Рост цепи РНК идет только в направлении 5'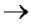 3'. Для начала синтеза РНК фермент не нуждается в полиили олигонуклеотидной затравке. Первый нуклеотид в РНК всегда пурин в форме трифосфата.
3'. Для начала синтеза РНК фермент не нуждается в полиили олигонуклеотидной затравке. Первый нуклеотид в РНК всегда пурин в форме трифосфата.
РНК-полимераза Е.coli - белок с четвертичной структурой. Одновременно в клетке присутствует около 7000 молекул РНК-полимеразы. Субъединичный состав
фермента: (2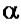 )
)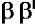
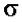 - holo-фермент (полный фермент). Без
- holo-фермент (полный фермент). Без 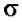 -фактора это core-
-фактора это core-
фермент (2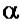 )
)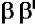 .
. 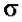 (сигма) -
(сигма) -
фактор - сменный фактор специфичности. Только holo-
фермент обладает высоким сродством к специфической последовательности нуклеотидов
– промотору. Как только произошла инициация транскрипции, 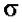 -фактор отделяется. Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
-фактор отделяется. Стадии узнавания и связывания, а также инициации осуществляются holo-ферментом. Элонгация и терминация осуществляются core-ферментом.
РНК-полимераза узнает промотор, покрывая 40-60 пар нуклеотидов - это AT-
богатые участки ДНК в положениях "-10" и "-35". Примерно 5% промоторов у прокариот имеют только участок "-10", однако хорошо узнаются РНК-полимеразой.
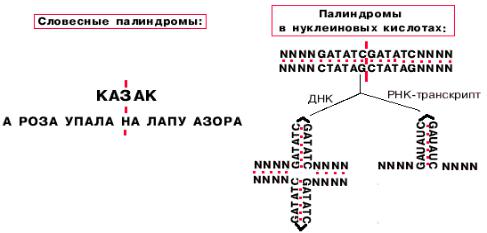
Такие промоторы представлены палиндромными последовательностями,
принимающими форму креста при суперспирализации кольцевых молекул ДНК.
20
Палиндромы -
последовательности,
которые читаются одинаково слева направо и справа налево. Палиндромы первого порядка имеют одну ось симметрии,
второго - две, третьего - три.
Этапы транскрипции
Инициация. Промотор содержит стартовый сигнал транкрипции ТАТА-бокс – определенную последовательность нуклеотидов ДНК, присоединяющий инициирующий ТАТА-фактор. Этот ТАТА-фактор обеспечивает присоединение РНК-полимеразы к той нити ДНК, которая будет использоваться в качестве шаблона для транскрипции. Так как промотор ассиметричен, то он связывает РНК-полимеразу только в одной ориентации, что определяет направление транскрипции от 5’-конца к
3’-концу (5’ → 3’). Другие факторы инициации раскручивают спираль ДНК перед РНК-полимеразой. После синтеза затравочного фрагмента РНК длиной 8-10
рибонуклеотидов σ-фактор отрывается от фермента.
Элонгация. Белковые факторы элонгации обеспечивают продвижение РНК-
полимеразы вдоль ДНК и расплетание нитей ДНК на протяжении примерно 17
нуклеотидных пар. РНК-полимераза продвигается со скоростью примерно 40-50
нуклеотидов в секунду в направлении 5’ → 3’. Используя одновременно в качестве субстрата и источника энергии АТФ, ГТФ, ЦТФ, УТФ.
Терминация. РНК-полимераза остановится, когда достигнет терминирующих кодонов. С помощью белкового фактора терминации, так называемого ρ-фактора
(греч. ρ - "ро"), от матрицы ДНК отделяются фермент и синтезированная молекула РНК, которая является первичным транскриптом, предшественником мРНК или тРНК или рРНК.