

Матричные биосинтезы №1 Репликация. Репарация. Транскрипция
.pdf1
Тема занятия: «Матричные биосинтезы»
Репликация. Репарация. Транскрипция
1. Репликация – синтез ДНК: матрица, затравка, субстраты, кофактор,
ферменты и белки репликации
2.Основные повреждения в ДНК и их репарация
3.Транскрипция – синтез РНК: субстраты, кофактор. РНК-полимераза.
Обратная транскрипция
4. Процессинг РНК: посттранскрипционные превращения различных типов
РНК
5. Генетический код и его свойства. Значение тРНК в декодировании генетической информации

2
Открытия в молекулярной биологии
1944г. Доказательство генетической роли ДНК. Освальд Эйвери, Колин Мак-
Леод, Маклин Мак-Карти
1953г. Установление структуры ДНК. Джеймс Уотсон, Френсис Крик
1961г. Открытие генетической регуляции синтеза ферментов. Андре Львов,
Франсуа Жакоб, Жак Моно
1962г. Расшифровка генетического кода. Маршалл Нирнберг, Генрих Маттеи,
Северо Очоа
1967г. Синтез in vitro биологически активной ДНК. Артур Корнберг
(неформальный лидер молекулярной биологии)
1970г. Химический синтез гена. Гобинд Корана
1970г. Открытие фермента обратной транскриптазы и явления обратной транскрипции. Говард Темин, Дэвид Балтимор, Ренато Дульбеко
1974г. Открытие рестриктаз. Гамильтон Смит, Даниэль Натанс, Вернер Арбер
1978г. Открытие сплайсинга. Филипп Шарп
1982г. Открытие автосплайсинга. Томас Чек
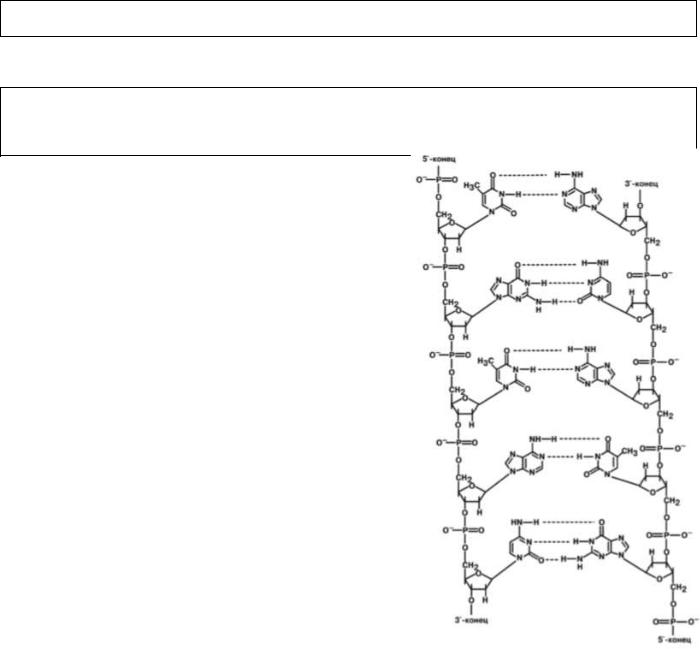
Основной фигурой матричных биосинтезов являются нуклеиновые кислоты.
Они представляют собой полимерные молекулы, в состав которых входят азотистые основания пяти типов, пентозы двух типов и остатки фосфорной кислоты. Азотистые основания в нуклеиновых кислотах могут быть пуриновыми (аденин, гуанин) и
пиримидиновыми (цитозин, урацил, тимин). В зависимости от строения углевода выделяют рибонуклеиновые кислоты – содержат рибозу (РНК), и
дезоксирибонуклеиновые кислоты – содержат дезоксирибозу (ДНК).
3
Центральная догма молекулярной биологии
В подавляющем большинстве случаев передача наследственной информации от материнской клетки к дочерней осуществляется при помощи ДНК. Для использования генетической информации самой клеткой необходимы РНК,
образуемые на матрице ДНК. Далее РНК непосредственно участвуют на всех этапах синтеза белковых молекул, обеспечивающих структуру и деятельность клетки. На вышесказанном основана центральная догма молекулярной биологии, согласно которой перенос генетической информации осуществляется только от нуклеиновой кислоты (ДНК и РНК). Получателем информации может быть другая нуклеиновая кислота (ДНК или РНК) и белок.
перенос генетической информации в направлении ДНК → РНК → белок
Правила Чаргаффа
в ДНК всегда А/Т=1; Г/Ц=1; (Г+Ц) / (А+Т) = К,
коэффициент специфичности, постоянен для каждого вида
ДНК – наиболее важная часть хромосом:
две двухцепочечные молекулы ДНК образуют одну хромосому. Наиболее хорошо хромосомы видны перед митозом и во время его. В
покоящихся клетках хромосомный материал выглядит нечетко и распределен по всему ядру.
В таком состоянии он получил название
"хроматин". В составе хроматина выделяют 60%
белка (гистоны и кислые белки), 35% ДНК и около 5% РНК.
4
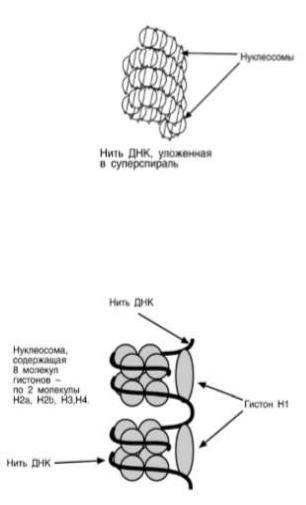
Хроматин уложен в виде сферических частиц – нуклеосом, соединенных друг с другом нитью ДНК. Нуклеосома представляет собой комплекс участка молекулы ДНК и восьми молекул гистонов. В составе нуклеосомы находятся по 2 молекулы гистонов Н2α, Н2β, Н3, Н4. Нить ДНК последовательно контактируя с гистонами Н2α, Н2β, Н4, Н3, Н3, Н4, Н2β, Н2α, наматывается на гистоновое ядро, которое
"маскирует" 146 пар оснований ДНК. Гистон Н1 связывается с нуклеосомой на участке входа и выхода ДНК, "склеивая" 2 оборота и "маскируя" еще 20 пар оснований. Всего замаскировано 166 пар оснований. Кроме нуклеосом, в ядре присутствуют еще 2 структуры: фибриллы диаметром 10 нм, состоящие из цепочки нуклеосом, и волокна, диаметром 30 нм, образующиеся при закручивании фибрилл в спираль. На виток спирали приходится 6-7 нуклеосом. Участок ДНК между нуклеосомами называется спейсерным (англ: space – пространство), его длина варьирует в зависимости от вида организма и типа клеток. У человека она составляет около 50 пар нуклеотидов. Благодаря наличию нуклеосом достигается уменьшение размеров хромосомы в 7 раз, далее происходит укладка в суперспираль и „суперсуперспираль".
Таким образом, благодаря гистонам размеры ДНК уменьшаются в тысячи раз: если длина ДНК достигает 6-
9 см (10-1), то размеры хромосом – всего несколько микрометров (10-6).
Хроматин может быть активным (эухроматин) и неактивным (гетерохроматин).
Активный хроматин содержит активные гены, т.е. те гены, с которых считывается информация. В активном хроматине нуклеосомная структура изменена или вообще отсутствует, благодаря чему ДНК становится доступной для соответствующих ферментов.
5
Принципы строения ДНК
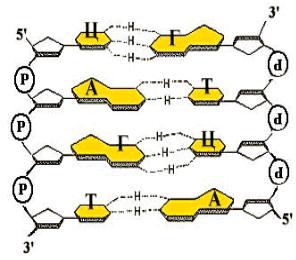
1.Нерегулярность. Существует регулярный сахаро-фосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
2.Антипараллельность. ДНК состоит из двух полинуклеотидных цепей,
ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой.
3. Комплементарность
(дополнительность). Каждому азотистому основанию одной цепи соответствует строго определенное азотистое основание другой цепи. Соответствие задается химией. Пурин и пиримидин в паре образуют водородные связи. В паре A-Т 2
водородные связи, в паре Г-Ц - 3.
4.Наличие регулярной вторичной структуры. Две комплементарные,
антипараллельно расположенные полинуклеотидные цепи образуют правую спираль
с общей осью.
Функции ДНК
1.ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода.
2.Воспроизведение и передача генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
3.Реализация генетической информации в виде белков, а также любых других соединений, образующихся с помощью белков-ферментов. Функция обеспечивается процессами транскрипции и трансляции.
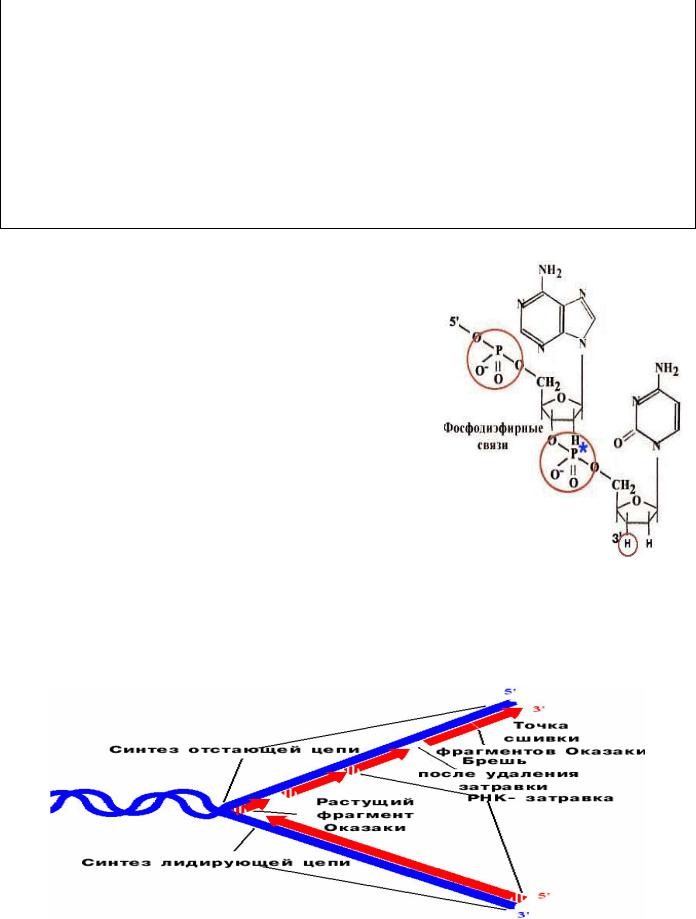
Строение РНК
Рибонуклеиновая кислота (РНК) представляет собой последовательность
рибонуклеозидмонофосфатов, связанных друг с другом 5’-3’-фосфодиэфирными

6
связями. РНК отличается от ДНК однонитевой структурой, наличием урацила вместо тимина и рибозы вместо дезоксирибозы. В клетке присутствует четыре типа РНК.
Рибосомальные РНК (рРНК) у прокариот и эукариот различны и отличаются величиной седиментации (S, величиной скорости оседания молекулы). У прокариот три разновидности рРНК: 5S, 16S и 23S. У эукариот четыре разновидности: 5S, 5,8S, 18S и 28S. Рибосомальные РНК участвуют в построении рибосом, внутриклеточных белоксинтезирующих органелл. Рибосомы состоят из двух неравных субчастиц,
малой и большой.
У прокариот малую (30S) субчастицу образуют белки, 23S-
рРНК и 5S-рРНК; большую (50S) – белки и 16S-рРНК. У
эукариот малую (40S) субчастицу образуют белки и 18S-рРНК,
большую (60S) – белки и 5S-, 5,8S-, 28S-рРНК.
Матричные РНК (мРНК) представляют собой линейную последовательность нуклеотидов. К 5’-концу молекулы присоединен метилгунозиндифосфат, на 3’-конце имеется полиадениловая последовательность. Их функция – информационная, т.е. перенос информации о структуре белков от ДНК к месту их синтеза.
Транспортные РНК (тРНК) бактерий и эукариот
включают 73-93 нуклеотида. Они переносят аминокислоты из цитозоля к рибосомам. Вторичная структура тРНК напоминает клеверный лист, а третичная
– латинскую букву L. В «клеверном листе» выделяют четыре участка (или ветви, петли), каждый из которых имеет собственную функцию: антикодоновый,
псевдоуридиловый,

7
дигидроуридиловый, акцепторный. На 5’-конце тРНК находится гуаниловый
нуклеотид, на 3’-конце – триплет Ц-Ц-А.
Малые РНК – используются для созревания мРНК и некоторых других
клеточных процессов.
|
Виды РНК |
|
|
Виды РНК |
Размер в нуклеотидах |
|
|
|
|
gРНК - геномные РНК |
10000-100000 |
|
|
|
|
mРНК - матричные РНК |
100-100000 |
|
|
|
|
tPHK - транспортные РНК |
70-90 |
|
|
|
|
rРНК - рибосомные РНК |
несколько классов от 100 до 500000 |
|
|
|
|
sРНК - малые РНК |
100-300 |
|
|
|
|
Отличия между ДНК и РНК
|
|
|
|
Параметры |
ДНК |
РНК |
|
|
|
|
|
|
|
|
|
Сахар |
Дезоксирибоза |
Рибоза |
|
|
|
|
|
|
|
|
|
Азотистые основания |
А, Т, Г, Ц |
А, У, Г, Ц |
|
|
|
|
|
|
|
|
|
Количество цепей в |
99,99% двойная спираль |
99,99% одноцепочечная |
|
молекуле |
0,01% одноцепочечная |
0,01% двухцепочечная |
|
|
|
|
|
|
|
|
|
Форма |
Все одноцепочечныекольцевые. |
|
|
Большинство двухцепочечных - |
Линейные молекулы |
||
молекулы |
|||
линейные, частькольцевые. |
|
||
|
|
||
|
|
|
|
|
|
|
1. Репликация – синтез ДНК: матрица, затравка, субстраты, кофактор,
ферменты и белки репликации
Репликация ДНК - процесс, осуществляемый комплексом ферментов и
белков, образующих идентичные копии ДНК для передачи генетической информации
в поколениях клеток и организмов.
8
Синтез ДНК в клетке происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы: митоз (М), синтетическую (S),
пресинтетическую (G1, от англ. gap - интервал), постсинтетическую (G2). Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой
35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины
– это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК).
Активные комплексы циклин-ЦЗК фосфорилируют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Принципы репликации:
1.Комплементарность.
2.Антипараллельность.
3.Униполярность.
4.Полуконсервативность
5.Потребность в затравке.
6.Прерывистость.
Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи и всегда в направлении 5'
3'.
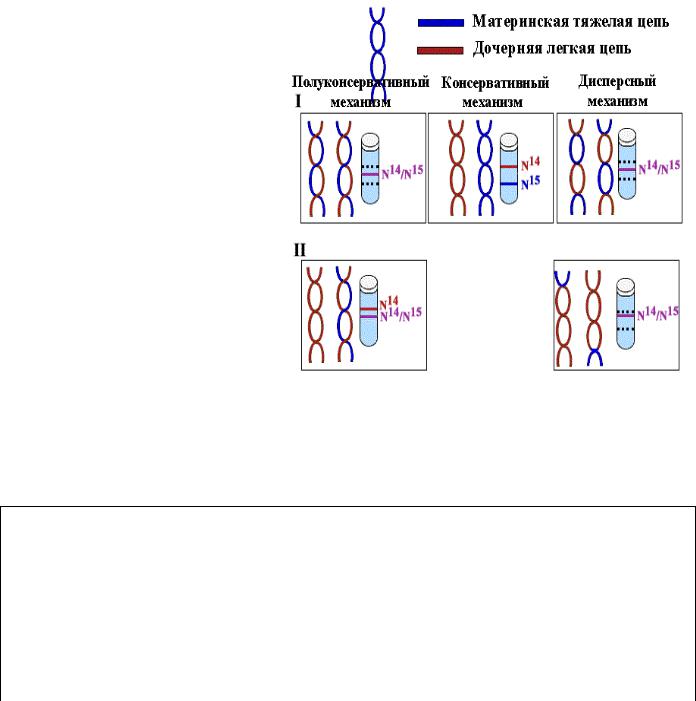
Полуконсервативность означает, что каждая дочерняя ДНК состоит из одной матричной цепи и одной вновь синтезированной.
Доказательство полуконсервативного характера репликации
Для выяснения вопроса о характере расхождения цепей по дочерним молекулам Мэтт Мезельсон и Фрэнк Сталь в 1958 г. разработали метод равновесного центрифугирования в градиенте плотности CsCl для разделения ДНК по удельной плотности. E. сoli выращивали на протяжении нескольких поколений на среде,
содержащей тяжелый изотоп азота (N15) для того, чтобы вся ДНК была "тяжелой".
9
Перед очередным раундом деления клетки в среде заменяли N15 на легкий изотоп N14
с тем, чтобы вновь синтезированные цепи были "легкими". После репликации ДНК выделяли и центрифугировали в градиенте плотности CsCl. Равное распределение
"тяжелых" и "легких" цепей между всеми молекулами исключало возможность консервативного способа, согласно которому одна дочерняя клетка получает материнскую ДНК, а другая - вновь синтезированную, обе цепи которой являются новыми.
Матрицей для синтеза новых цепей служит одноцепочечная ДНК.
Затравкой является 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей. Доказательством того, что затравка - 3'-
гидроксильный конец, является эксперимент с дидезоксинуклеозидтрифосфатом. Если такой активированный нуклеотид сделать меченым по α-
фосфату, то он включается в растущую полимерную цепь и всегда обнаруживается на ее 3'-конце. Это
говорит о том, что он сам включается, но дальнейший рост цепи невозможен, т.к. нет
3'-гидроксильного конца. Это доказывает и униполярность репликации в направлении 5' 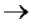 3'.
3'.
Прерывистость репликации происходит в виде фрагментов Оказаки.
10
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла.
Механизм репликации, как установили эксперименты Мэтью Мезельсон и Франклин Сталь в 1958 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия. Весь процесс репликации идет в S-фазу клеточного цикла, в то время, когда клетка готовится к делению. Как матричный биосинтез,
репликация требует наличия нескольких условий:
•матрица – в ее роли выступает материнская ДНК;
•растущая цепь – дочерняя ДНК;
•субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ;
•источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ;
•ферменты.
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin - начало). На каждой ДНК млекопитающих точек ori насчитывается около 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием "репликативных пузырей". В каждом таком "пузыре" имеются две "репликативные вилки", в которых происходит расплетание, раскручивание и непосредственный синтез ДНК. Репликативные вилки удаляются друг от друга. В
целом вся репликация ДНК у эукариот заканчивается за 9 часов.
В каждой репликативной вилке идет синтез ДНК в направлении от 5'-конца к
3'-концу, т.е. 5'-конец новой ДНК остается свободным, следующие нуклеотиды присоединяются к 3'-гидроксильной группе предыдущего нуклеотида. Поскольку нити ДНК антипараллельны, то непрерывно синтезируется только одна нить, а
именно та, на которой направление движения репликативной вилки совпадает с направлением 3' → 5'.