

Введение в биохимию гормонов
.pdfТема: «Введение в биохимию гормонов»
План:
1.Сигнальные молекулы - гормоны: определение, классификация по: химическому строению, биологическому действию, характеристика.
2.Рецепторы: определение, характеристика, классификация.
3.Понятие о G-белках, общая схема передачи сигнала через рецепторы, сопряженные с G-белками.
4.Трансмембранная передача сигнала с участием систем:
аденилатциклазной, инозитолтрифосфатной.
5.Передача сигнала через рецепторы, обладающие ферментативной активностью.
6.Передача сигнала через ядерные рецепторы (цитозольный механизм).
Клеточная сигнализация мембран
Гормоны (греч. hormao – привожу в движение) – это вещества,
вырабатываемые специализированными клетками и регулирующие обмен веществ в отдельных органах и во всем организме в целом. Для всех гормонов характерна большая специфичность действия и высокая биологическая
активность.
Гормоны можно классифицировать по химическому строению,
растворимости, локализации их рецепторов и влиянию на обмен веществ.
|
КЛАССИФИКАЦИЯ ПО СТРОЕНИЮ |
|
|
Гормоны – |
|
Стероидные |
|
|
|||
производные |
Пептидные гормоны |
||
гормоны |
|||
аминокислот |
|
||
|
|
||
Адреналин |
Адренокортикотропный гормон (АКТГ) |
Кортизол |
|
Норадреналин |
Соматотропный гормон (СТГ) |
Альдостерон |
|
Тироксин |
Тиреотропный гормон (ТТГ) |
Эстрадиол |
|
Трийодтиронин |
Лактотропный гормон (пролактин, ПЛГ) |
Прогестерон |
|
|
Лютеинизирующий гормон (ЛГ) |
Тестостерон |
|
|
Фолликулостимулирующий гормон (ФСГ) |
Кальцитриол |
|
|
Меланоцитстимулирующий гормон (МСГ) |
|
|
|
Антидиуретический гормон (АДГ, вазопрессин) |
|
|
|
Окситоцин |
|
|
|
Кальцитонин |
|
Паратгормон
Инсулин
Глюкагон
КЛАССИФИКАЦИЯ ГОРМОНОВ ПО ВЛИЯНИЮ НА ОБМЕН ВЕЩЕСТВ
КЛАССИФИКАЦИЯ ПО МЕСТУ СИНТЕЗА
Место синтеза |
Гормоны |
|
|
|
|
|
Кортиколиберин, тиреолиберин, гонадолиберин, сома- |
|
Гипоталамус |
толиберин, меланолиберин. |
|
|
Пролактостатин, соматостатин, меланостатин. |
|
Гипофиз |
СТГ, АКТГ, ЛТГ, ТТГ, АДГ, МСГ, ФСГ, ЛГ, оксито- |
|
цин, |
||
|
||
|
Инсулин, глюкагон, кортизол, тироксин, адреналин, |
|
Периферические железы |
альдостерон, эстрадиол, эстриол, тестостерон, кальци- |
|
|
тонин, паратгормон, кальцитриол |
М Е Х А Н И З М Ы Д Е Й С Т В И Я Г О Р М О Н О В Для регуляции деятельности клетки с помощью гормонов, находящихся
в плазме крови, необходимо обеспечить возможность клетки воспринимать и обрабатывать этот сигнал. Эта задача усложняется тем, что сигнальные молекулы (нейромедиаторы, гормоны, эйкозаноиды) имеют разную химическую природу, реакция клеток на сигналы должна быть различной по направленности и адекватной по величине.
В связи с этим, эволюционно сформировались два основных механизма действия сигнальных молекул по локализации рецептора:
1.Мембранный – рецептор расположен на мембране.
2.Цитозольный – рецептор расположен в цитозоле.
МЕ М Б Р А Н Н Ы Е Р Е Ц Е П Т О Р Ы
Взависимости от способа передачи гормонального сигнала в клетку
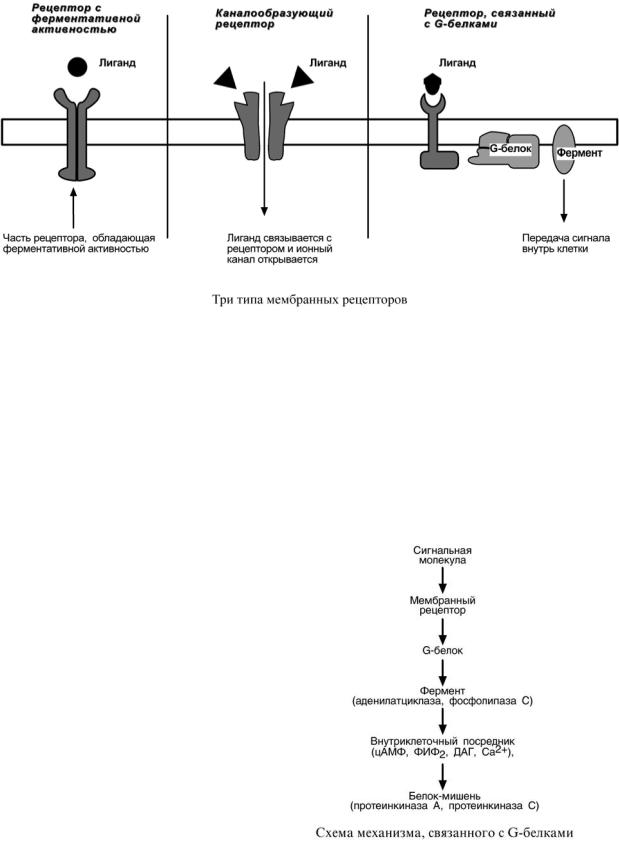
выделяют три класса мембраносвязанных рецепторов:
1. Рецепторы, обладающие каталитической активностью – при
взаимодействии лиганда с рецептором активируется внутриклеточная часть
(домен) рецептора, имеющий тирозинкиназную или тирозинфосфатазную
или гуанилатциклазную активность. По этому механизму действуют СТГ,
инсулин, пролактин, интерлейкины, ростовые факторы, интерфероны
α, β, γ.
2.Каналообразующие рецепторы –
присоединение лиганда к рецептору вызывает открытие ионного канала на мембране. Таким образом, действуют нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин, гистамин, глутамат);
3.Рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве G-белка. G- белок влияет на ферменты, образующие вторичные посредники (мессенджеры). Последние передают сигнал на внутриклеточные белки.
Кпоследнему классу относится аденилатциклазный и кальцийфосфолипидный механизмы.
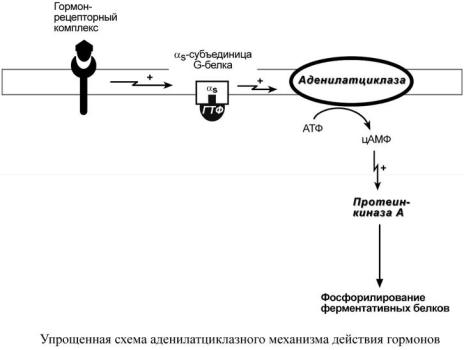
Аденилатциклазный механизм
По аденилатциклазному механизму действуют гормоны гипофиза (ТТГ,
ЛГ, МСГ, ФСГ. АКТГ), кальцитонин, соматостатин, глюкагон,
паратгормон, адреналин (через α2- и β-адренорецепторы), вазопрессин
(через V2-рецепторы).
1. Взаимодействие лиганда с рецептором приводит к изменению конформации последнего.
2. Это изменение передается на G-белок, который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ.
В составе G-белков α-субъединицы бывают двух типов по отношению к ферменту: активирующие αs и ингибирующие αi.
3.В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно ГДФ заменяется на ГТФ.
4.Активированная таким образом αS-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ.
5.цАМФ – вторичный мессенджер – в свою очередь, взаимодействует
спротеинкиназой А и активирует ее.
6.Протеинкиназа А фосфорилирует ряд ферментов, среди которых
киназа фосфорилазы гликогена, гликогенсинтаза, ТАГ-липаза.
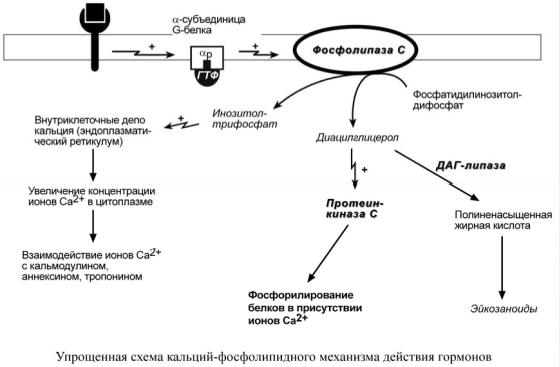
Кальций-фосфолипидный механизм
По этому механизму действуют вазопрессин (через V1-рецепторы),
адреналин (через α1-адренорецепторы), ангиотензин II.
Принцип работы этого механизма совпадает с предыдущим, но вместо аденилатциклазы мишеневым ферментом для α-субъединицы служит фосфолипаза С. Фосфолипаза С расщепляет мембранный фосфолипид фосфатидилинозитолдифосфат до вторичных мессенджеров
инозитолтрифосфата (ИФ3) и диацилглицерола (ДАГ).
Инозитолтрифосфат открывает кальциевые каналы в эндоплазматическом ретикулуме, что вызывает увеличение концентрации ионов Cа2+ в цитоплазме и активацию определенных
кальций-связывающих белков (например, кальмодулина).
Диацилглицерол совместно с ионами Са2+ активирует протеинкиназу С,
участвующую в процессах клеточной пролиферации. Кроме этого,
диацилглицерол имеет и другую сигнальную функцию: он может распадаться на 1-моноацилглицерол и полиеновую кислоту (обычно
арахидоновую), из которой образуются эйкозаноиды.
Гуанилатциклазный механизм
Данный механизм пока изучен недостаточно подробно, но известно, что по нему работает атриопептин (предсердный натрийуретический пептид),
оксид азота (NO) и другие, пока малоизученные, соединения.
Гуанилатциклазный механизм передачи сигнала в общих чертах схож с аденилатциклазным: после появления какой-либо сигнальной молекулы сигнал воспринимается ферментом гуанилатциклазой и далее передается при помощи вторичного мессенджера цГМФ. Он воздействует на протеинкиназу
G, которая фосфорилирует определенные белки, чем изменяет активность клетки.
В отличие от аденилатциклазы, данный фермент имеет четыре разновидности, три из которых связаны с мембраной, четвертый – цитозольный. Мембраносвязанные формы гуанилатциклазы работают как рецепторы, обладающие каталитической активностью.
Цитозольный фермент является димером и содержит в своем составе гем, он непосредственно взаимодействует в цитозоле с сигнальной молекулой (например,оксид азота).
Ц И Т О З О Л Ь Н Ы Е Р Е Ц Е П Т О Р Ы
По цитозольному механизму действуют стероидные гормоны и гормоны щитовидной железы. Будучи гидрофобными молекулами, они попадают в клетки путем простой диффузии и прочно связываются с рецепторами,
расположенными в цитоплазме. Комплекс гормонрецептор в некоторых случаях дополнительно модифицируется и активируется. Далее он проникает в ядро, где может связываться с ядерным рецептором. В результате гормон приобретает сродство к ДНК. Связываясь с гормон-чувствительным элементом в ДНК, гормон влияет на транскрипцию определенных генов и изменяет концентрацию РНК в клетке и соответственно количество определенных белков в клетке.

Иерархия гормональных систем
Большинство гормональных систем взаимосвязаны между собой и образуют иерархическую лестницу. Верхнюю часть лестницы занимает система гормонов гипоталамуса, контролируемая центральной нервной системой. На стимулирующие или тормозящие стимулы из ЦНС клетки гипоталамуса отвечают выбросом стимулирующих или ингибирующих веществ, которые носят название либерины или статины соответственно. Эти нейрогормоны с кровотоком достигают аденогипофиза, где стимулируют (либерины) или ингибируют (статины) биосинтез и секрецию тропных гормонов.
Тропные гормоны воздействуют на периферические железы, стимулируя выделение соответствующих гормонов. К подобным системам относятся группы гормонов тиреоидной функции, глюкокортикоидной функции и профиль половых гормонов.
Регуляция таких систем осуществляется по принципу обратной отрицательной связи:
накопление гормонов периферических желез тормозит секрецию рилизинг-факторов
гипоталамуса и тропных гормонов гипофиза. Наиболее значительно это проявляется в отношении регуляции стероидных гормонов.
Подавляющее действие на активность эндокринных желез может оказывать и конечный ответ клеток-мишеней.