

Структура и функции биологических мембран
.pdfТема: «Структура и функции биологических мембран»
План:
1.Мембраны клетки: функции, строение, свойства.
2.Липидный состав мембран, роль в формировании липидного
бислоя.
3.Белки мембран - интегральные, поверхностные, «заякоренные».
4.Механизмы переноса веществ через мембраны: простая диффузия, первичноактивный транспорт (Na+-K+-АТФ-аза), симпорт и антипорт, вторичноактивный транспорт (глю/ Na+).
СТРУКТУРА И ФУНКЦИИ БИОЛОГИЧЕСКИХ МЕМБРАН
1.Функции мембран клеток
Биологические мембраны – это клеточные структуры,
ограничивающие цитоплазму клетки от окружающей среды и внутриклеточных органелл. Внутри клетки мембраны также образуют единую систему канальцев и замкнутых полостей. Мембрана - важнейший орган клетки, регулирующий взаимодействие как внутри клетки, так и клетки с окружающей средой. Если функции мембраны нарушаются, то происходит изменение нормального функционирования клеток и, как следствие, заболевание организма.
Основные функции мембран
•Барьерная - отделение клетки от окружающей среды и формирование внутриклеточных компарментов (отсеков);
•Транспортная - контроль и регулирование транспорта огромного разнообразия веществ через мембраны;
•Информационно-сигнальная - участие в обеспечении межклеточных взаимодействий, передаче внутрь клетки сигналов;
•Каталитическая

Мембраны органелл эукариотических клеток уникальны по своему составу и по характеру выполняемых функций.
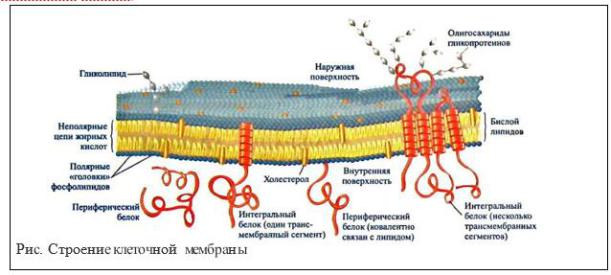
2. Общая схема строения биологических мембран
Биологические мембраны представляют собой «ансамбли» липидных и белковых молекул, удерживаемых вместе с помощью нековалентных взаимодействий.
Основу мембраны составляет двойной липидный слой, в
формировании которого участвуют фосфолипиды и гликолипиды.
Липидный бислой образован двумя рядами липидов, гидрофобные радикалы которых спрятаны внутрь, а гидрофильные группы обращены наружу и контактируют с водной средой. Белковые молекулы как бы «растворены» в
липидном бислое.
Липиды мембран. Мембранные липиды – амфифильные молекулы, т.е.
в молекуле есть как гидрофильные группы (полярные «головки»), так и алифатические радикалы (гидрофобные «хвосты»), самопроизвольно формирующие бислой. В большинстве эукариотических клеток они составляют около 30-70% массы мембраны. В мембранах присутствуют липиды трёх главных типов: фосфолипиды, гликолипиды и холестерин.
Все фосфолипиды можно разделить на 2 группы -
глицерофосфолипиды и сфингофосфолипиды.
Глицерофосфолипиды относят к производным фосфатидной кислоты. Наиболее распространённые глицерофосфолипиды мембран -
фосфатидилхолины и фосфатидилэтаноламины. Каждый глицерофосфолипид, например фосфатидилхолин, представлен несколькими десятками фосфатидилхолинов, отличающихся друг от друга строением жирнокислотных остатков. На долю инозитолфосфатов прихо-
дится всего 2-8%. Инозитол в составе фосфатидилинозитолов может быть фосфорилирован по С4 (фосфатидилинозитол-4-монофосфат) или С4 и С5 (фосфатидилинозитол- 4,5-бисфосфат).

Рис. Глицерофосфолипиды мембран
Специфические фосфолипиды внутренней мембраны митохондрий -
кардиолипины (дифосфатидилглицеролы), построенные на основе глицерола и двух остатков фосфатидной кислоты. В плазматических мембранах клеток в значительных количествах содержатся сфингомиелины:
Рис.Структура кардиолипинов

Сфингомиелины построены на основе церамида – ацилированного ами-
носпирта сфингозина Полярная группа состоит из остатка фосфорной кислоты и холина, этаноламина или серина. Сфингомиелины - главные липиды миелиновой оболочки нервных волокон:
Рис. 29 Строение сфингомиелина
Гликолипиды. В гликолипидах гидрофобная часть представлена церамидом.
Гидрофильная группа – углеводный остаток, присоединённый гликозидной связью к гидроксильной группе у первого углеродного атома церамида. В
зависимости от длины и строения углеводной части различают цереброзиды,
содержащие моноили олигосахаридный остаток, и ганлиозиды к ОН-группе которых присоединён сложный, разветвлённый олигосахарид, содержащий N-
ацетилнейраминовую кислоту:
Рис. 28 Структура цереброзида
ствах гликолипиды содержатся в мембранах клеток мозга, эритроцитов,
эпителиальных клеток. Ганглиозиды эритроцитов различаются строением олигосахаридных цепей, проявляющих антигенные свойства.
Холестерол присутствует во всех мембранах животных клеток. В составе

мембран растений холестерола нет. Его молекула состоит из жёсткого гидрофобного ядра и гибкой углеводородной цепи, единственная гидроксильная группа является «полярной головкой»
Функции мембранных липидов. Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций.
1) Липиды формируют среду для функционирования мембранных белков,
принимающих в ней нативную конформацию.
2) Некоторые мембранные липиды - предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфатидилинозитол-4,5-
бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАГ), активатора протеинкиназы С и инозитол-1,4,5-
трифосфата (ИФ3) - регулятора кальциевого обмена в клетке ДАГ, ИФ3,
протеинкиназа С и Са2+ – участники инозитолфосфатной системы передачи сигнала.
3)Некоторые липиды выполняют «якорную» функцию, например к фосфатидилинозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности клетки
4)Липиды могут быть аллостерическими активаторами мембранных
ферментов. Например, β-гидроксибутиратдегидрогеназа, участвующая в
окислении кетоновых тел, локализована на внутренней мембране митохондрий.
Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина. Креатинкиназа, фермент, катализирующий образование макроэргического соединения креатинфосфата. Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны
митохондрий.
4.Свойства биологических мембран
4.1Трансмембранная асимметрия липидов. Каждая мембрана клетки замкнута,
т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам - эту особенность мембран называют трансмембранной
(поперечной) асимметрией.
Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными «головками» стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную
«головку», больше.
Фосфатидилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем.
4.2 Жидкокристаллическое состояние. Молекулы в мембране размещены не беспорядочно, в их расположении наблюдается дальний порядок.
Фосфолипидные молекулы находятся в двойном слое, а их гидрофобные хвосты приблизительно параллельны друг другу. Есть порядок и в ориентации полярных гидрофильных головок. Физическое состояние, при котором есть дальний порядок во взаимной ориентации и расположении молекул, но агрегатное состояние жидкое, называется жидкокристаллическим состоянием.
Мембранные липиды могут находиться в нескольких фазовых состояниях:
кристаллическое и жидкокристаллическое. Различаются они плотностью упаковки и подвижностью находящихся в бислое белковых молекул. При более плотной упаковке ацильные цепи липидов расположены под углом, близким к 90°, и все С—С-связи находятся в транс-конформации (максимально вытянуты). Фазовый переход приводит к увеличению подвижности ацильных цепей в бислое, увеличению угла их наклона и уменьшению плотности упаковки. Латеральная подвижность мембранных белков после фазового перехода возрастает, увеличивается вероятность образования их ассоциатов.
Жидкокристаллические структуры очень чувствительны к изменению температуры. В мембранных фосфолипидах при понижении температуры

происходит переход из жидкокристаллического в гель состояние. При этом изменяется взаимное положение гидрофобных хвостов и увеличивается толщина двойного слоя.
При переходе в гель-состояние в бислое образуются сквозные каналы,
радиусом 1-3 нм, по которым через мембрану могут переноситься ионы и низкомолекулярные вещества.
4.3 Жидкостность мембран. Для мембран характерна жидкостность (те-
кучесть), способность липидов и белков к латеральной диффузии.
липидных молекул затрудняют полярные «головки», поэтому липиды,
находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость трансмембранной миграции по сравнению с липидами наружной стороны мембраны, мигрирующих медленнее или вообще не совершающими
«флип-флоп» перескоки.
Скорость перемещения молекул зависит от микровязкости мембран, которая, в
свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов.

Эти «изломы» препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно, и более «текучей». На текучесть мембран также влияют размеры углеводородных «хвостов» липидов, с
увеличением длины которых мембрана становится более «текучей».
бранах уменьшает подвижность жирных кислот (снижает текучесть мембран),
снижает латеральную диффузию липидов и белков и поэтому может влиять на функции мембранных белков.
4.4.Самосборка мембран. В водном растворе происходит самосборка мембран

и замыкание мембран с образованием липидных пузырьков называемых липосомами. Это свойство мембран обусловлено амфифильностью молекул фосфолипидов, составляющих основу бислоя.
Белки мембран
Поверхностные белки - прикрепляются к мембране разными способами. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счет гидрофобных, электростатических или других нековалентных сил. Периферические белки, легко вымываются из мембран растворами солей или даже дистиллированной водой. Такие белки способны обратимо связываться с бислоем и часто совершают челночные перемещения между мембраной и ее окружением (например, апобелки липопротеионов).
Деление мембранных белков на периферические и интегральные определяется их структурой, количеством гидрофобных аминокислот и их расположением в бислое фосфолипидов.
Периферические белки отличаются от интегральных меньшей глубиной проникновения в бислой и более слабыми белок-липидными взаимодействиями