

Регуляторные системы организма. Биохимия стероидных гормонов
.pdf5.Системы передачи гормонального сигнала в клетки-мишени:
аденилатциклазная, инозитолфосфатная, гуанилатциклазная.
РЕЦЕПТОРЫ, СВЯЗАННЫЕ С G-БЕЛКАМИ
Аденилатциклазный механизм
По аденилатциклазному механизму действуют гормоны гипофиза (ТТГ, ЛГ,
МСГ, ФСГ. АКТГ), кальцитонин, соматостатин, глюкагон, паратгормон,
адреналин (через α2- и β-адренорецепторы), вазопрессин (через V2-рецепторы).
1.Взаимодействие лиганда с рецептором приводит к изменению конформации последнего.
2.Это изменение передается на G-
белок, который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ. В составе G-белков α-
субъединицы бывают двух типов по отношению к ферменту: активирующие
αs и ингибирующие αi.
3. В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно в αS-
субъединице ГДФ заменяется на ГТФ. 4. Активированная таким образом
αS-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ.
5. Активность αS-субъединицы сохраняется до тех пор, пока ее ГТФ-
азный центр не гидролизует ГТФ до ГДФ. После этого происходит быстрая ресборка G-белка, все возвращается в исходное состояние.
11

6. цАМФ – вторичный мессенджер – в свою очередь связывается с регуляторными субъединицами протеинкиназы А (R) и вызывает их диссоциацию от каталитических. В результате каталитические субъединицы (C)
становятся активными.
7. Активная протеинкиназа А фосфорилирует белки и ряд ферментов,
среди которых киназа фосфорилазы гликогена, гормон-чувствительная липаза,
транскрипционный фактор CREB (cAMP-response element-binding protein).
Кальций-фосфолипидный механизм
По этому механизму действуют вазопрессин (через V1-рецепторы), адреналин
(через α1-адренорецепторы), ангиотензин II. Принцип работы этого механизма совпадает с предыдущим, но вместо аденилатциклазы мишеневым ферментом для α-субъединицы служит фосфолипаза С:
12
1.Взаимодействие лиганда с рецептором приводит к изменению конформации последнего.
2.Это изменение передается на G-белок, который состоит из трех субъединиц (α, β и γ), α-субъединица связана с ГДФ.
3.В результате взаимодействия с рецептором β- и γ-субъединицы отщепляются, одновременно в αp-субъединице ГДФ заменяется на ГТФ.
4.Активированная таким образом αp-субъединица стимулирует фосфолипазу С, которая гидролизует фосфатидилинозитолдифосфат (ФИФ2)
вторичных мессенджеров диацилглицерола (ДАГ) и инозитолтрифосфата (ИФ3). 5. Активность αp-субъединицы сохраняется до тех пор, пока ее ГТФ-азный центр не гидролизует ГТФ до ГДФ. После этого происходит быстрая ресборка
G-белка, все возвращается в исходное состояние.
6. ИФ3 – вторичный мессенджер – в свою очередь открывает кальциевые каналы в эндоплазматическом ретикулуме, что вызывает увеличение концентрации ионов Cа2+ в цитоплазме и активацию определенных кальций-
связывающих белков, меняя их активность (например, кальмодулина).
7. Диацилглицерол является активатором протеинкиназы С, которая в присутсвии ионов Ca2+ фосфорилирует свои белковые субстраты.
Протеинкиназа С в основном участвует в процессах клеточной пролиферации.
Кроме этого, диацилглицерол имеет и другую сигнальную функцию: он может распадаться на 1-моноацилглицерол и полиеновую кислоту (чаще арахидоновую), из которой образуются эйкозаноиды.
13

Гуанилатциклазный механизм
Данный механизм передачи сигнала в общих чертах схож с аденилатциклазным: после появления какой-либо сигнальной молекулы сигнал воспринимается ферментом гуанилатциклазой и далее передается при помощи вторичного мессенджера цГМФ. Последний воздействует на протеинкиназу G,
которая фосфорилирует определенные белки, чем изменяет активность клетки.
Гуанилатциклаза присутствует во многих органах (сердце, сетчатка глаза,
легкие, почки, надпочечники, эндотелий кишечника) и активно участвует в регуляции внутриклеточного метаболизма. В отличие от аденилатциклазы, этот фермент имеет четыре разновидности, три из которых связаны с мембраной,
четвертый – цитозольный. Соотношения этих двух форм фермента в различных тканях разные. Мембраносвязанные формы гуанилатциклазы работают как рецепторы, обладающие каталитической активностью. Их лигандами являются
14

предсердный (ANP) и мозговой (BNP) натрийуретические пептиды. Действие этих пептидов на гладкие мышцы сосудов вызывает их расслабление и снижение артериального давления, в канальцах почек – подавляет реабсорбцию ионов Na+.
Цитозольная (растворимая) форма фермента взаимодействует в цитозоле с другими сигнальными молекулами – оксид азота, пероксинитрит, супероксид-
анион-радикал, которые связываются с гемом активного центра фермента и изменяют его активность. Эти лиганды выступают как вазодилататоры,
ингибиторы агрегации тромбоцитов, противовоспалительные агенты.
ЦИТОЗОЛЬНЫЕ РЕЦЕПТОРЫ
По цитозольному механизму действуют стероидные гормоны и гормоны щитовидной железы. Будучи гидрофобными молекулами, они попадают в клетки путем простой диффузии и прочно связываются с рецепторами,
расположенными в цитоплазме. Комплекс гормон-рецептор в некоторых случаях дополнительно модифицируется и активируется. Далее он проникает в ядро, где может связываться с ядерным рецептором. В результате гормон приобретает сродство к ДНК. Связываясь с гормон-чувствительным элементом в ДНК,
гормон влияет на транскрипцию определенных генов и изменяет концентрацию РНК в клетке и, соответственно, количество определенных белков в клетке.
15
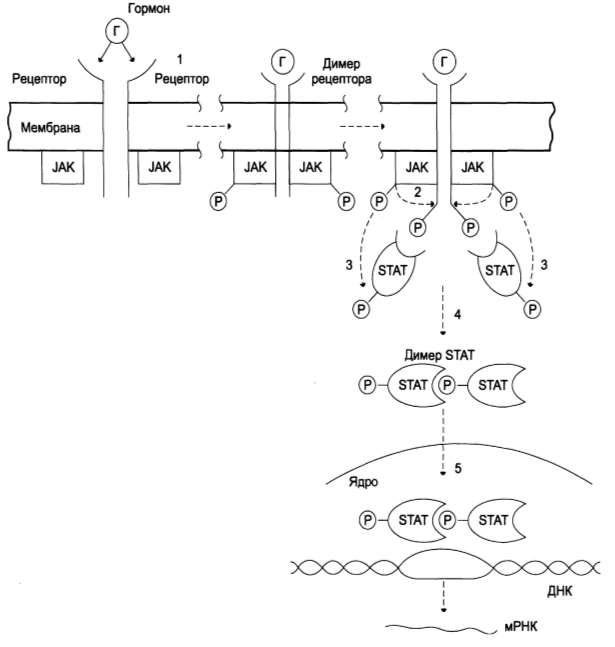
Рис. Механизм передачи сигнала через мембранные рецепторы,
ассоциированные с Янус-киназами (JAK)
1 – гормон взаимодействует с мембранным рецептором и вызывает димеризацию рецептора. Янус-киназы (цитоплазматические тирозинкиназы, имеющие два активных центра) связываются с димером мембранного рецептора, что приводит к их активации и аутофосфорилированию
2 – Янус-киназы (JAK) фосфорилируют димер рецептора по остаткам тирозина
3 – комплекс фосфорилированного димера рецептора с Янус-киназами связывает особые цитоплазматические белки (STAT), которые фосфорилируются Янус-
киназами
16

4 – фосфорилированные белки STAT активируются, образуя димер
5 – димер STAT перемещается из цитозоля в ядро, связывается с промоторным участком ДНК и индуцирует транскрипцию генов
6.Стероидные гормоны: классификация, общая схема биосинтеза,
катаболизм.
Стероидные гормоны являются производными холестерола.
Женские гормоны: эстрогены синтезируются в фолликулах яичников,
прогестерон – в желтом теле. Частично гормоны могут образовываться в адипоцитах в результате ароматизации андрогенов.
Мужские гормоны: на 95% тестостерон синтезируется в клетках Лейдига семенников и на 5% в периферических тканях при метаболизме андрогенов,
образованных в сетчатой и пучковой зонах надпочечников. Механизм действия Цитозольный.
17
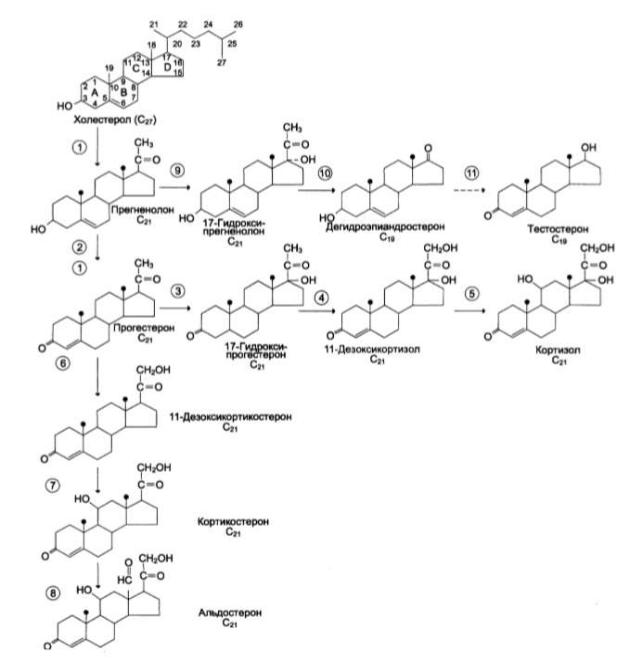
Рис. Строение и основные этапы синтеза кортикостероидов:
1 – превращение холестерола в прегненолон (гидрокислаза, отщепляющая боковую цепь)
2 – образование прогестерона (3-β-гидроксистероиддегидрогеназа)
3, 4, 5 – реакции синтеза кортизола (3 – 17-гидроксилаза, 4 – 21гидроксилаза,
5 – 11гидроксилаза)
6, 7, 8 – путь синтеза альдостерона (6 - 21гидроксилаза, 7- 11гидроксилаза,
8 - 18гидроксилаза, 18 - гидроксидегидрогеназа)
18
9, 10, 11 – путь синтеза тестостерона (9 - 17гидроксилаза, 10 – 17,20-лиаза, 11 –
дегидрогеназа)
Катаболизм гормонов коры надпочечников происходит в печени.
Здесь протекают реакции гидроксилирования, окисления и восстановления гормонов. Продукты катаболизма кортикостероидов (кроме кортикостерона и альдостерона) выводятся мочой в форме 17-кетостероидов, образующихся в результате отщепления боковой цепи. Эти продукты метаболизма выделяются преимущественно в виде конъюгатов с глюкуроновой и серной кислотами. 17-
окси- и 17-кетостероиды образуются также при катаболизме половых гормонов,
которые имеют у С 17 гидкроксиили кетогруппы. У мужчин 2/3 кетостероидов образуется за счет тестостерона (12-17 мг/сут). У женщин 17-кетостероиды образуются преимущественно за счет кортикостероидов (7-12 мг/сут).
Определение 17-кетостероидов в моче позволяет оценить, как количество глюкокортикоидов, секретируемых корой надпочечеников, так и функцию надпочечников.
7.Гормоны коры надпочечников – глюкокортикоиды: химическая природа,
структура, регуляция биосинтеза (гипоталамо-гипофизарно-
надпочечниковая ось), механизм действия, транспорт, клетки-мишени,
влияние на обмен веществ
ГЛЮКОКОРТИКОИДЫ. Строение
Глюкокортикоиды являются производными холестерола – стероидные гормоны.
Основным гормоном у человека является кортизол.
Синтез Осуществляется в сетчатой и пучковой зонах коры надпочечников
(схему синтеза кортизола см выше).
Регуляция синтеза и секреции Активируют: АКТГ, обеспечивающий нарастание концентрации кортизола в утренние часы, к концу дня содержание
19

кортизола снова снижается. Кроме этого, имеется нервная стимуляция секреции гормонов. Уменьшают: кортизол по механизму обратной отрицательной связи.
Механизм действия цитозольный.
Рис. Внутриклеточная локализация синтеза кортизола
1 - аденилатциклазный комплекс, 2 – холестеролэстераза, 3 – протеинкиназа А,
4 – холестеролдесмолаза отщепляет боковую цепь холестерола ХС – холестерол, ЭХС – эфиры холестерола
20