

Методичка по биохимии. 1 курс - 2 модуль
.pdfРис.1 Обмен фруктозы в печени
2.Наследственные нарушения обмена фруктозы.
Врожденные аномалии обмена фруктозы могут быть связаны с генетическими дефектами ферментов фруктокиназы и фрутозо-1- фосфатальдолазы.
Существует доброкачественная эссенциальная фруктоземия,
встречается с частотой 1:130 000 и протекающей безо всяких отрицательных симптомов.Связанасврожденнымнедостаткомферментафруктокиназы,т.е. в организме не образуется фруктозо-1-фосфат. В результате обмен фруктозы возможен только путем фосфорилирования до фруктозо-6-фосфата, но эта реакция тормозится глюкозой, вследствие чего фруктоза накапливается в крови. «Почечный порог» для фруктозы очень низок, поэтому фруктозурия обнаруживается уже при концентрации фруктозы в крови 0,73 ммоль/л.
Наследственная непереносимость фруктозы, возникающая при генетически обусловленном дефекте фруктозо-1-фосфатальдолазы, не проявляется, пока ребёнок питается грудным молоком, т.е. пока пища не содержитфруктозы.Симптомывозникают,когдаврациондобавляютфрукты, соки, сахарозу. Рвота, боли в животе, диарея, гипогликемия и даже кома и судороги возникают через 30 мин после приёма пищи, содержащей фруктозу. У маленьких детей и подростков, продолжающих принимать фруктозу, развиваются хронические нарушения функций печени и почек.
Дефект альдолазы фруктозо-1-фосфата сопровождается накоплением фруктозо-1-фосфата, который ингибирует активность фосфоглюкомутазы, превращающей глюкозо-1-фосфат в глюкозо-6-фосфат и обеспечивающей включение продукта гликогенфосфорилазной реакции в метаболизм. Поэтому происходит торможение распада гликогена на стадии образования глюкозо-1- фосфата, в результате чего развивается гипогликемия. Как следствие, ускоряется мобилизация липидов и окисление жирных кислот. Следствием ускорения окисления жирных кислот и синтеза кетоновых тел, замещающих энергетическую функцию глюкозы, может быть метаболический ацидоз, так как кетоновые тела являются кислотами и при высоких концентрациях снижают рН крови.
Лечение фруктоземии заключается в исключении из рациона соков, ягод, фруктов и овощей. При своевременном выключении фруктозы ребенок развивается нормально и проявления заболевания приобретают обратное развитие.
3.Обмен галактозы в печени. Галактоземия.
Галактоза сначала подвергается фосфорилированию по 1-му атому углерода. Отличием от обмена фруктозы является превращение в глюкозу не напрямую, а через синтез УДФ-галактозы, которая впоследствии изомеризуется в глюкозу.
Галактоземия возникает при нарушении обмена галактозы, обусловленном наследственным дефектом любого из трёх ферментов, включающих галактозу в метаболизм глюкоз:
галактокиназы, частота дефекта 1:500000,
галактозо-1-фосфат-уридилтрансферазы, частота дефекта
1:40000,
эпимеразы, частота дефекта реже 1:1000000.
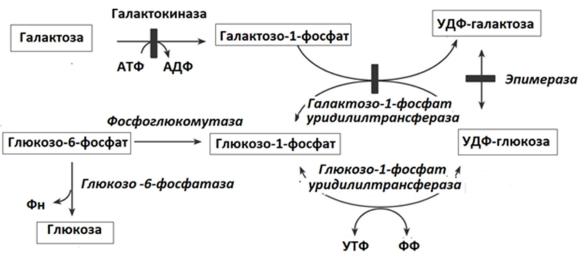
Рис.2 Обмен галактозы в печени
Это заболевание проявляется очень рано, и особенно опасно для детей, так какосновным источникомуглеводов для них служит материнское молоко, содержащее лактозу.
Патогенез. Избыток галактозы превращается в спирт галактитол (дульцит), накапливающийся в хрусталике и осмотически привлекающий сюда воду. Изменяется солевой состав, нарушается конформация белков хрусталика,чтоприводитккатарактевмолодомвозрасте.Катарактавозможна даже у плодов матерей с галактоземией, употреблявших молоко во время беременности.
При дефекте галактозо-1-фосфатуридилтрансферазы АТФ постоянно расходуется на фосфорилирование галактозы и дефицит энергии угнетает активность многих ферментов, "токсически" действуя на нейроны, гепатоциты, нефроциты. Как результат возможны задержка психомоторного развития, умственная отсталость, некроз гепатоцитов и цирроз печени.
Ранние симптомы галактоземии: рвота, диарея, дегидратация,
уменьшениемассытела,желтуха.Онипоявляютсявскорепослерождения,как толькоребёнокначинаетполучатьмолоко.Вкрови,мочеитканяхповышается концентрация галактозы.
Основы лечения. Исключение из рациона молока и других источников галактозы позволяет предотвратить развитие патологических симптомов. Однако сохранностьинтеллектаможетбыть достигнута только приранней, не позднее первых 2 месяцев жизни, диагностике и вовремя начатом лечении.
4. Пути нарушения обмена углеводов в организме человека: гипер- и гипогликемия: причины, компенсация.
Гипергликемическим является состояние, при котором концентрация глюкозы в крови более 6 ммоль/л.
По происхождению выделяют две группы таких состояний: 1. Физиологические
алиментарные – связаны с приемом пищи и продолжаются в норме не более 2 часов после еды.
нейрогенные – нервное напряжение, стимулирующее секрецию адреналина и мобилизацию гликогена в печени,
гипергликемия беременных – связана с относительной недостаточностьюинсулинаприувеличениимассытелаипотребностью
плода в глюкозе.
2. Патологические:
Сахарный диабет
при заболеваниях гипофиза, коры и мозгового слоя
дпочечников, щитовидной железы, поджелудочной железы, при органических поражениях ЦНС.
5. Глюкозотолерантный тест
Глюкозотолерантный тест (ГТТ) — лабораторный метод исследования, применяемый в эндокринологии для диагностики нарушения толерантности к глюкозе (предиабет) и сахарного диабета.
Обследуемый принимает раствор глюкозы из расчёта 1 г на 1 кг массы тела (сахарная нагрузка).
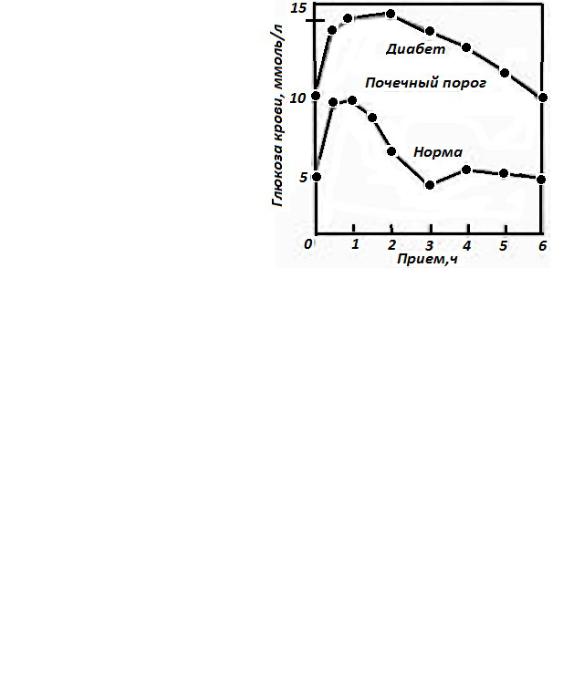
Концентрацию глюкозы в крови измеряют в течение 2-3 ч с интервалами в 30 мин.
У здорового человека уровень глюкозы в крови растет, а затем быстро снижается.
Уровень глюкозы в плазме крови выше 10 ммоль/л через 2 ч после сахарной нагрузки свидетельствует о сахарном диабете;
Рис.48 Сахарная кривая в норме и при сахарном диабете
Рекомендуемая литература по дисциплине «Молекулярная медицина»:
1.Северин Е.С., Биохимия [Электронный ресурс]: учебник / под ред. Е. С. Северина. - 5-е изд., испр. и доп. - М.: ГЭОТАР-Медиа, 2016. - 768 с. - ISBN 978-5-9704-3762-9 - Режим
доступа: http://www.studmedlib.ru/book/ISBN9785970437629.html
2.Авдеева Л.В., Биохимия [Электронный ресурс] / Авдеева Л.В., Алейникова Т.Л., Андрианова Л.Е., Белушкина Н.Н., Волкова Н.П., Воробьева С.А., Голенченко В.А., Губарева А.Е., Корлякова О.В., Лихачева Н.В., Павлова Н.А., Рубцова Г.В., Силаева С.А., Силуянова С.Н., Титова Т.А. - М. : ГЭОТАР-
Медиа, 2014. - 752 с. - ISBN 978-5-9704-3043-9 - Режим доступа: http://www.studmedlib.ru/book/ISBN9785970430439.html
3.Губарева А.Е., Биологическая химия. Ситуационные задачи и тесты [Электронный ресурс]: учеб. пособие / А. Е. Губарева [и др.]; под ред. А. Е.
Губаревой. - М.: ГЭОТАР-Медиа, 2016. - 528 с. - ISBN 978-5-9704-3561-8 -
Режим доступа: http://www.studmedlib.ru/book/ISBN9785970435618.html
Тема: «Обмен гликогена, регуляция, нарушения»
План:
1.Биосинтез гликогена (гликогеногенез): схема процесса, ферменты.
2.Распад гликогена (гликогенолиз): схема процесса, ферменты. Особенности распада гликогена в печени и скелетных мышцах.
3.Регуляция обмена гликогена.
4.Гликогеновые болезни: гликогенозы и агликогенозы.
1. Биосинтез гликогена (гликогеногенез)
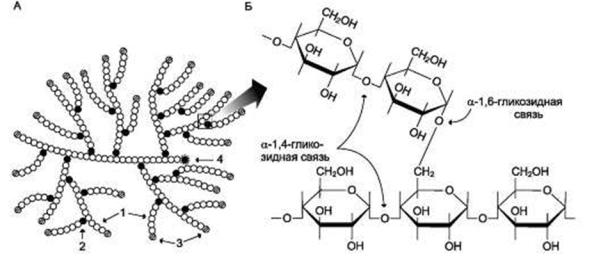
Гликоген представляет собой разветвленный гомополимер глюкозы, в
котором остатки глюкозы соединены в линейных участках α-1,4-
гликозидными связями, а в точках ветвления – α-1,6- гликозидными связями.
Эти связи образуются примерно с каждым десятым остатком глюкозы,
то есть точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Гликоген иногда называется животным крахмалом,
так как его структура похожа на амилопектин - компонент растительного крахмала. В отличие от крахмала, гликоген имеет более разветвленную и компактную структуру.
Рис.1 Строение молекулы гликогена
Биологическая роль гликогена - основной резервный полисахарид в животных тканях, из которого по мере метаболической потребности могут высвобождаться глюкозо-6-фосфат или глюкоза. Запас углеводов в виде
гликогена обусловлен тем, что гликоген как ВМС в отличие от глюкозы не повышает осмотического давления клеток.
Локализация биосинтеза гликогена. Гликоген синтезируется в период пищеварения (абсорбтивный период: 1-2 часа после приема углеводной пищи)
в основном в печени и в мышцах. 2/5 запасов гликогена (примерно 150 грамм)
откладывается в паренхиме печени в виде глыбок (запас гликогена в печени может составлять примерно 5% от её массы). Остальной гликоген откладывается в мышцах и других органах. В мышцах запасается около 1%
гликогена, однако масса мышечной ткани значительно больше и поэтому общее количество гликогена в мышцах в 2 раза больше, чем в печени.
Гликоген может синтезироваться во многих клетках, например в нейронах,
макрофагах, клетках жировой ткани, но содержание его в этих тканях незначительно. В организме может содержаться до 450 г гликогена.
Этапы биосинтеза гликогена. Синтез гликогена сложный,
многоступенчатый процесс, который включает 3 стадии:
1.образование активной формы глюкозы;
2.синтез линейной цепочки гликогена, т.е. образование
α-1,4-гликозидных связей
3.ветвление линейной цепочки гликогена, т.е.
образование α-1,6-гликозидных связей.
Процесс синтеза гликогена требует затрат энергии, так включение одного мономера в полисахаридную цепь сопряжено с расходованием АТФ и УТФ.
В метаболические пути глюкоза и другие моносахариды включаются только в виде фосфорных эфиров. Фосфорилирование свободных моносахаридов - обязательная реакция на пути их использования в клетках,
она приводит к образованию более реакционноспособных соединений и поэтому может рассматриваться как реакция активации. Эту реакцию во многих тканях катализирует фермент гексокиназа, а в печени и поджелудочной железе – глюкокиназа. Фосфорилирование глюкозы –
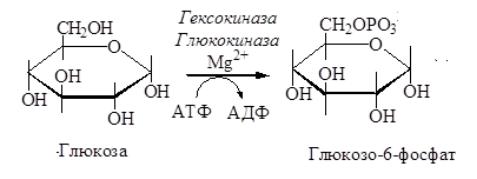
практически необратимая реакция. Образование глюкозо-6-фосфата в клетке
– это своеобразная «ловушка» для глюкозы, так как мембрана клетки непроницаема для фосфорилированной глюкозы:
Глюкокиназа имеет высокое значение Км = 10 ммоль/л и катализирует фосфорилирование глюкозы в гепатоцитах в период пищеварения
(абсорбтивный период). В этот период концентрация глюкозы в воротной вене больше, чем в других отделах кровяного русла, и может превышать 10
ммоль/л. Максимальная активность глюкокиназы обеспечивает поступление глюкозы в клетки печени и ее фосфорилирование. Глюкокиназа, в отличие от гексокиназы, не ингибируется продуктом реакции - глюкозо-6-фосфатом.
Гексокиназа отличается от глюкокиназы высоким сродством к глюкозе и низким значением Км <0,1ммоль/л. Следовательно, этот фермент, в отличие от глюкокиназы, активен при концентрации глюкозы в крови,
соответствующей физиологической норме, и обеспечивает потребление глюкозы мозгом, эритроцитами и другими тканями между приемами пищи
(постабсорбтивный период).
Рис. 2 Образование глюкозо-6-фосфата
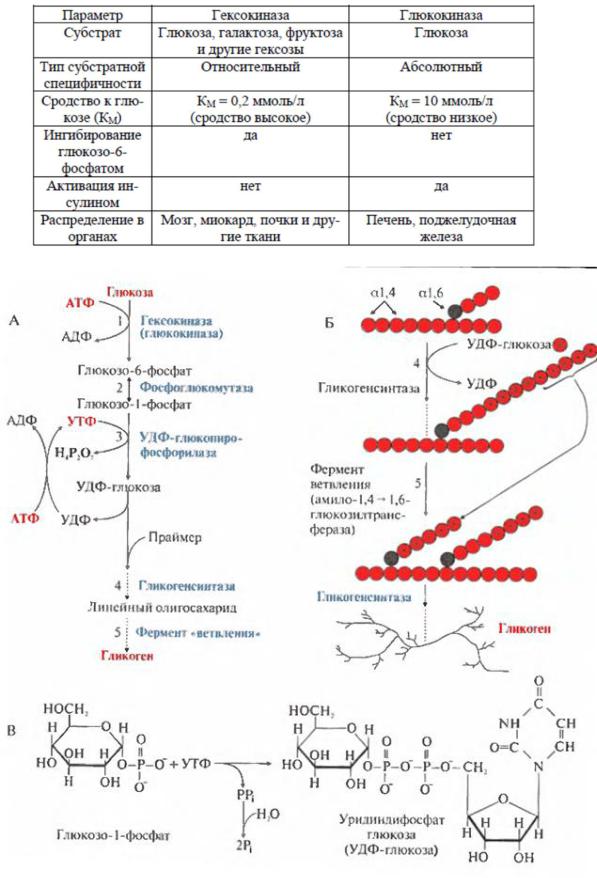
Таблица 1
Характеристика ферментов глюкокиназы и гексокиназы:
Рис. 3 Синтез гликогена
А – синтез гликогена (общая схема); Б – полимеризация и ветвление молекулы гликогена В – образование УДФглюкозы.
1 – фосфорилирование глюкозы с использованием молекулы АТФ и образование глюкозо-6-фосфата;
2– превращение глюкозо-6– фосфата в глюкозо-1-фосфат.
3– образование УДФ-глюкозы – субстрата для синтеза гликогена с использованием молекулы УТФ;
4– перенос глюкозного остатка на праймер – «затравку» (цепь из 4-8 остатков глюкозы с участием гликогенсинтазы, которая удлиняет праймер, соединяя остатки глюкозы α-1,4-гликозидными связями; 5 – образование разветвлений в молекуле гликогена: фермент ветвления - амило- 1,4-1,6-
гликозилтрансфераза образует боковую цепь путем переноса концевого олигосахарида из 5-6 остатков на внутренний остаток глюкозы этой же или другой цепи, образуя точку ветвления, где концевой остаток глюкозы соединяется с олигосахаридом α-1,6- гликозидной связью. Таким образом, образуется точка ветвления в молекуле.
Таким образом:
1) синтез гликогена требует много энергии - для присоединения 1
молекулы глюкозы к фрагменту гликогена используется 1молекула АТФ и 1
молекула УТФ;
2)для инициации процесса обязательно наличие затравки гликогена;
3)в образовании зрелой молекулы гликогена принимают участие гликогенсинтаза и амило- 1,4-1,6-гликозилтрансфераза; 4) этот процесс не безграничен – избыток глюкозы превращается в липиды.
2. Распад гликогена (гликогенолиз)
Мобилизация (распад) гликогена происходит в интервалах между приемами пищи (постабсорбтивный период) и ускоряется во время физической работы. Этот процесс осуществляется путем последовательного отщепления остатков глюкозы, в виде глюкозо-1-фосфата с помощью гликогенфосфорилазы, расщепляющей α-1,4-гликозидные связи. Этот фермент не расщепляет α 1,6-гликозидные связи в местах разветвлений,
поэтому необходимы еще два фермента, после действия которых остаток глюкозы в точке ветвления освобождается в форме свободной глюкозы
.Сначала три оставшихся до точки ветвления глюкозных остатка переносятся