

Методичка по биохимии. 1 курс - 2 модуль
.pdfПри включении глицерол-3-фосфата в глюконеогенез происходит его дегидрирование НАД-зависимой дегидрогеназой с образованием дигидроксиацетонфосфата, который далее превращается в глюкозу.
Суммарная реакция синтеза глюкозы из глицерола
2Глицерол+2АТФ+2НАД+2Н2О→глюкоза+2АДФ+2Н3РО4+2НАДН+Н
Синтез глюкозы из лактата. Использование лактата связано с его превращением в печени в пируват (цикл Кори). Лактат как источник пирувата важен не столько при голодании, сколько при нормальной жизнедеятельности организма.
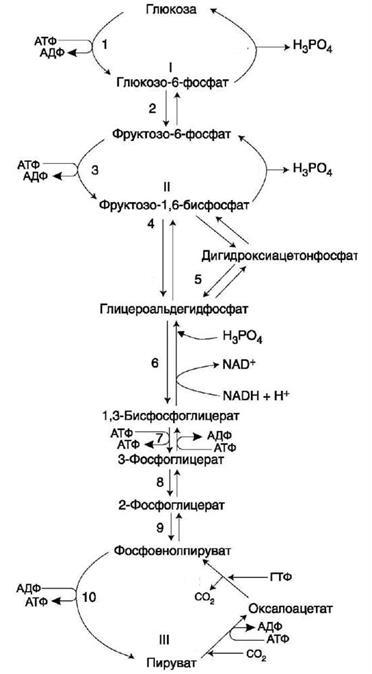
Рис. 5 Общая схема гликолиза и глюконеогенеза
4. Регуляция процессов гликолиза и глюконеогенеза
Процессы гликолиза и глюконеогенеза имеют разное биологическое значение, поэтому скорость каждого из них определяется физиологическим состоянием организма.
Гликолитическое окисление глюкозы как процесс, ведущий к выработке энергии, активируется при недостатке энергии в клетке. Глюконеогенез необходим при гипогликемии и как любой анаболический процесс, наоборот,
идет с затратой энергии.
Превращения метаболитов в глюконеогенезе и гликолизе скоординированы так, что один процесс активен, а другой нет. Если одновременно активны оба процесса, то следует гидролиз четырех молекул нуклеозидтрифосфатов (2 АТФ и 2 ГТФ) в каждом цикле. Оба процесса экзергоничны, поэтому термодинамически невыгодна одновременная их активация.
Переключение метаболизма печени с гликолиза на глюконеогенез и наоборот происходит при помощи:
•- аллостерических механизмов регуляции активности ключевых
ферментов:
•- ковалентной модификации ферментов путем фосфорилирования
(дефосфорилирования) с участием инсулина и глюкагона;
• - индукции (репрессии) синтеза ключевых ферментов,
катализирующих реакции субстратных или «холостых» циклов.
Регуляторные ферменты гликолиза - гексокиназа, глюкокиназа,
фосфофруктокиназа и пируваткиназа.
Регуляторные ферменты глюконкогенеза: глюкозо-6-фосфатаза,
фруктозо-1,6-дифосфатаза (фосфатаза фруктозо-1,6-бифосфата), ФЕП-
карбоксикиназа и пируваткарбоксилаза);
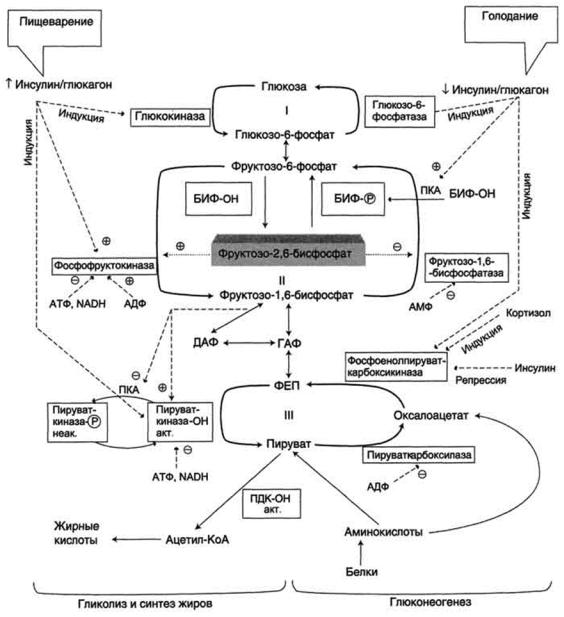
Регуляция гликолиза и глюконеогенеза в печени направлена на необратимые стадии гликолиза и глюконеогенеза, которые формируют три субстратных цикла. Эти циклы обозначены цифрами I, II, III на рис.5. Название
«субстратный цикл» означает объединение реакций синтеза и распада субстрата. Название «холостой» отражает результат работы подобного цикла,
заключающийся в бесполезном расходовании АТФ. Хотя существование
«холостых» циклов нелогично, тем не менее они могут функционировать.
Более того, эти циклы могут быть мишенью регуляторных воздействий, так как составляющие их реакции катализируют разные ферменты. Реципрокное изменение активности этих ферментов предотвращает одновременное протекание противоположных процессов.
Изменение в печени гликолитического направления на глюконеогенез и обратно при смене абсорбтивного состояния на постабсорбтивное или при голодании происходит главным образом в результате регуляции активности ферментов, катализирующих реакции субстратных циклов.
Направление реакций первого субстратного цикла регулируется главным образом концентрацией глюкозы.
При пищеварении (абсорбтивный период) концентрация глюкозы в крови повышается (до 10 ммоль/л). Активность глюкокиназы в этих условиях максимальна. Вследствие этого ускоряется реакция Глюкоза → Глюкозо-6-
фосфат. Поскольку глюкокиназа печени не ингибируется глюкозо-6-
фосфатом (в отличие от гексокиназы мышц), то основная часть глюкозо-6-
фосфата направляется в гликолиз и на синтез гликогена.
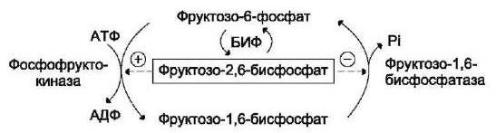
Направление реакций второго субстратного цикла зависит от активности фосфофруктокиназы и фруктозо-1,6-бисфосфатзы. Регулирует активность этих ферментов. фруктозо-2,6-бисфосфат,. который одновременно выполняет функцию аллостерического активатора фосфофруктокиназы и аллостерического ингибитора фруктозо-1,6-бисфосфатзы.
Рис. 6 Регуляция реакций II субстратного цикла фруктозо-2,6-бисфосфатом.
Фруктозо-2,6-бисфосфат образуется в абсорбтивный период путем фосфорилирования фруктозо-6-фосфата при участии бифункционального
фермента (БИФ).
Образование фруктозо-2,6-бисфосфата требует затрат АТФ, а при образовании фруктозо-6-фосфата из фруктозо-2,6-бисфосфата высвобождается неорганический фосфат.
БИФ катализирует реакции превращения фруктозо-6-фосфата во фруктозо-2,6-бисфосфат. БИФ может находиться в двух формах:
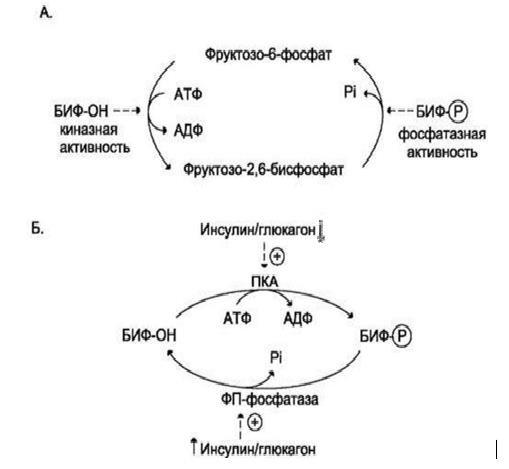
дефосфорилированной (БИФ-ОН) и
фосфорилированной (БИФ-ОРО3Н2).
БИФ-ОН обладает киназной активностью и способствует образованию фруктозо-2,6-бисфосфата. БИФ-ОРО3Н2 обладает фосфатазной активностью и способствует уменьшению концентрации фруктозо-2,6-бисфосфата и образованию фруктозо-6-фосфата.
Наличие двух активностей у БИФ определило название фермента -
«бифункциональный». БИФ присутствует только в гепатоцитах.
Киназную и фосфатазную реакции катализируют разные активные центры БИФ, которые функционируют по очереди в зависимости от состояния фермента фосфорилированного или дефосфорилированного.
Фосфорилирование и дефосфорилирование БИФ связано с ритмом
питания и регулируется гормонами.
Рис.7 (А). Реакции, катализируемые бифункциональным ферментом (БИФ) в печени (Б) Регуляция активности БИФ
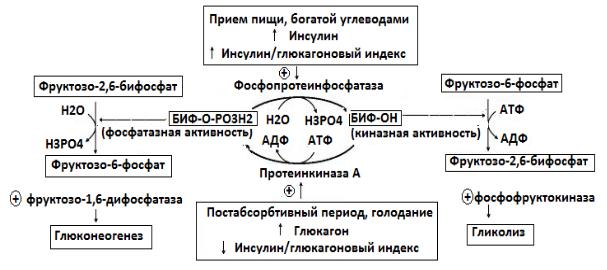
Инсулин в абсорбтивный период пищеварения активирует фермент фосфопротеинфосфатазу, что приводит к появлению формы БИФ-ОН,
активации фосфофруктокиназы и усилению гликолиза.
Глюкагон в постабсорбтивный период) активирует аденилатциклазную систему, что вызывает активацию протеинкиназы А и появление формы БИФ-
ОРО3Н2.
Результатом уменьшения количества фруктозо-2,6-бисфосфата в постабсорбтивном периоде будет снижение активности фосфофруктокиназы,
замедление гликолиза и переключение гликолиза на глюконеогенез.
Рис.8 Общая схема регуляции реакций II субстратного цикла фруктозо- 2,6-бисфосфатом
Направление реакций третьего субстратного цикла зависит от активности пируваткиназы, фосфорилированная форма которой неактивна, а
дефосфорилированная активна.
Регуляция активности пируваткиназы в печени осуществляется путем фосфорилирования (дефосфорилирования) в зависимости от ритма питания.
Дефосфорилирование пируваткиназы происходит в период пищеварения,
когда инсулин активирует фосфопротеинфосфатазу, которая дефосфорилирует пируваткиназу, переводя ее в активное состояние.
Следовательно, реакция превращения фосфоенолпируват в пируват ускоряется при пищеварении (абсорбтивный период).
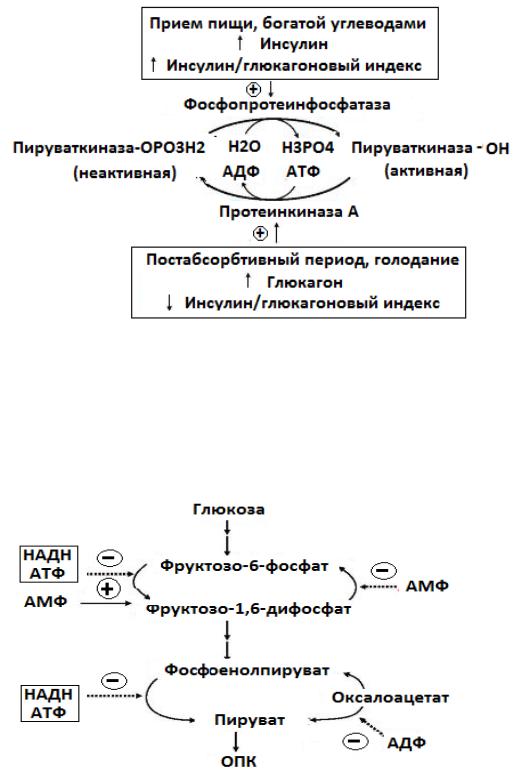
В постабсорбтивном состоянии пируваткиназа при действии глюкагона на печень переходит в фосфорилированное неактивное состояние.
Рис. 9 Регуляция пируваткиназы в печени.
Регуляция энергетического статуса гепатоцитов осуществляется
путем изменения скорости аэробного распада глюкозы. АТФ, AДФ, АМФ, а
также NADH, NAD+ служат аллостерическими эффекторами ключевых ферментов гликолиза и глюконеогенеза.
Рис.10 Регуляция аэробного гликолиза и глюконеогенеза в печени аллостерическим статусом клетки
При высокой концентрации АТФ и НAДH, характерных для высокоэнергетического статуса клетки, происходит ингибирование ключевых
ферментов гликолиза - фосфофруктокиназы и пируваткиназы, что вызывает торможение гликолиза. Высокая концентрация AMФ вызывает активацию фосфофруктокиназы (фермент гликолиза) и ингибирование фосфатазы фруктозо-1,6-бисфосфата (фермент глюконеогенеза), а AДФ ингибирует пируваткарбоксилазу, замедляя глюконеогенез. Таким образом, в случае низкого энергетического статуса клетки наблюдается активация гликолиза и ингибирование глюконеогенеза.
Индукция (репрессия) синтеза ключевых ферментов регулируется с
помощью гормонов.
Впериод пищеварения инсулин:
•индуцирует синтез глюкокиназы, фосфофруктокиназы и пируваткиназы, что приводит к активации гликолиза;
•вызывает репрессию фосфоенолпируваткарбоксикиназы и замедляет глюконеогенез.
Впостабсорбтивный период глюкагон повышает транскрипцию генов и синтез ключевых ферментов глюконеогенеза -
фосфоенолпируваткарбоксикиназы, фруктозо-1,6-бисфосфатазы и глюкозо-6-
фосфатазы, в результате чего активируется глюконеогенез.
Стероидные гормоны (кортизол) в период длительного голодания индукцию ферментов глюконеогенеза – пируваткарбоксилазы и фосфоенолпируваткарбоксикиназы.
Существенное влияние на глюконеогенез оказывает этанол.
Метаболизм этанола на 90% происходит в печени.
Превращение этанола включает две реакции дегидрирования с образованием ацетил-КоА и его последующее окисление в цитратном цикле.
Алкогольдегидрогеназа содержится в основном в печени (95%), а также в других органах (мозге, почках, легких, кишечнике). Для окисления суточной нормы углеводов (500 г) требуется такое же количество NAD+, как и для окисления 125 г этанола. Частично окисление этанола протекает под действием микросомальных ферментов окисления.
В результате катаболизма этанола увеличивается количество NADH, что приводит к смещению лактатдегидрогеназной реакции в сторону образования лактата, снижению образования пирувата и замедлению глюконеогенеза.
Рис.11 Общая схема метаболизма глюкозы в печени
Рекомендуемая литература по дисциплине «Биологическая химия»:
1.Северин Е.С., Биохимия [Электронный ресурс]: учебник / под ред. Е.
С. Северина. - 5-е изд., испр. и доп. - М.: ГЭОТАР-Медиа, 2016. - 768
с. - ISBN 978-5-9704-3762-9 - Режим
доступа: http://www.studmedlib.ru/book/ISBN9785970437629.html
2.Авдеева Л.В., Биохимия [Электронный ресурс] / Авдеева Л.В.,
Алейникова Т.Л., Андрианова Л.Е., Белушкина Н.Н., Волкова Н.П.,
Воробьева С.А., Голенченко В.А., Губарева А.Е., Корлякова О.В.,
Лихачева Н.В., Павлова Н.А., Рубцова Г.В., Силаева С.А., Силуянова С.Н., Титова Т.А. - М. : ГЭОТАР-Медиа, 2014. - 752 с. - ISBN 978-5- 9704-3043-9 - Режим
доступа: http://www.studmedlib.ru/book/ISBN9785970430439.html
3.Губарева А.Е., Биологическая химия. Ситуационные задачи и тесты
[Электронный ресурс]: учеб. пособие / А. Е. Губарева [и др.]; под ред.
А. Е. Губаревой. - М.: ГЭОТАР-Медиа, 2016. - 528 с. - ISBN 978-5- 9704-3561-8 - Режим
доступа: http://www.studmedlib.ru/book/ISBN9785970435618.html