

Методичка по биохимии. 1 курс - 2 модуль
.pdfРис. 7 Общая схема пентозофосфатного цикла
Особенности пентозофосфатного пути в различных тканях.
В отличие от др. основных путей метаболизма углеводов функционирование пентозофосфатного цикла нельзя представить в виде линейной последовательности реакций, приводящей непосредственно от 1
молекулы глюкозо-6-фосфата к 6 молекулам CO2. Пентозофосфатный цикл характеризуется возможностью многообразных взаимопревращений его метаболитов, происходящих по нескольким альтернативным путям.
Если клетка нуждается в больших количествах НАДФН, например,
синтез жирных кислот и холестерина в печени, то активно протекает ПФЦ:
идут реакции окислительной и неокислительной ветви, а образованные фруктозо-6-фосфат и глицеральдегидфосфат превращаются в глюкозо-6-
фосфат и цикл повторяется снова.
Если клетке нужны НАДФН и энергия АТФ (как в эритроцитах), то образованные фруктозо-6-фосфат и глицеральдегидфосфат включаются в гликолиз.
Если клетка растет и делится, то ей необходимы НАДФН и рибозо-5-
фосфат. В этом случае второй этап идти не будет, весь образованный рибулозо-5-фосфат превратится в рибозо-5-фосфат и используется на синтез нуклеотидов. НАДФН используется для синтеза дезоксирибонуклеотидов.
В тканях, которые не испытывают значительную потребность в
НАДФН, (например, скелетные мышцы) функционирует только неокислительная стадия ПФП, причем ее реакции идут в обратную сторону начиная с фруктозо-6-фосфата до рибозо-5-фосфат.
3. Регуляция пентозофосфатного пути
Неокислительные реакции могут начинаться от фруктозо-6-фосфата и завершаться образованием пентозофосфатов или они могут быть продолжением окислительных реакций, включая превращения пентозофосфатов в глюкозо-6-фосфат. Поток метаболитов через ПФП и образование НАДФ+ определяется функциональным состоянием клеток тканей.
Регуляция пентозофосфатного цикла осуществляется с помощью.
гормонов и аллостерически.
Регуляторным является первый фермент окислительного этапа пентозофосфатного пути глюкозо-6-фосфатдегидрогеназа, которая существует в фосфорилированной и дефосфорилированной форме. Активной является дефосфорилированная форма фермента, которая образуется под действием протеинфосфатазы, активируемой инсулином. В печени глюкозо-6-
фосфатдегидрогеназа, как и ключевые ферменты гликолиза и биосинтеза жирных кислот, индуцируется при увеличении соотношения инсулин/глюкагон после приёма богатой углеводами пищи, а снижается при голодании и сахарном диабете.
Аллостерически скорость ПФЦ регулируется в первую очередь
концентрацией НАДФН, который ингибирует глюкозо-6-
фосфатдегидрогеназу. Реакции окислительного пути протекают только в случае активного использования гидрированного НАДФН в восстановительных процессах или в системе антиоксидантной защиты.
Если клетки активно используют НАДФ+, то увеличивается концентрация НАДФ+, который повышает активность фермента и тем самым восполняет количество НАДФН.
Избыток рибозо-5-фосфата может превращаться в промежуточные продукты гликолиза.
3.1. Связь ПФП и гликолиза.
Пентозофосфатный путь и гликолиз тесно связаны и в зависимости от соотношения концентраций промежуточных продуктов, образовавшихся в клетке, возможны переключения с одного пути на другой.
При сбалансированной потребности в NADPH и пентозах в клетке
происходит окислительный путь синтеза пентоз. Если потребности в пентозах превышают потребности в NADPH, то окислительный путь шунтируется за счет использования метаболитов гликолиза: фруктозо-6-фосфат и глицероальдегидфосфат в реакциях неокислительного пути превращаются в пентозы.
Если же NADPH необходим в большей степени, чем пентозы, то
возможны два варианта:
-при высоком энергетическом статусе клетки излишки пентоз путем обратных реакций неокислительного пути превращаются в фруктозо-6-фосфат и глицероальдегидфосфат, из которых в процессе глюконеогенеза образуется глюкоза;
-при низком энергетическом статусе клетки из пентоз также образуются глицероальдегидфосфат и фруктозо-6-фосфат, которые затем включаются в гликолиз.
Глицеральдегид-3-фосфат в зависимости от условий и вида клеток может включаться в гликолиз, либо через диоксиацетонфосфат восстанавливаться в α-глицерофосфат и далее на синтез ТАГ. При необходимости из него могут получаться и гексозы.
4.Наследственные нарушения пентозофосфатного пути связаны
сгенетической недостаточностью фермента глюкозо-6-фосфатдегидрогеназы.
Следствием ферментного дефекта является снижение синтеза НАДФН в клетке. Особенно существенно это влияет на эритроциты, в которых окислительный этап пентозофосфатного цикла является единственным источником НАДФН. Из разнообразных функций НАДФН в данном случае имеет значение одна – участие в работе антиоксидантной системы.
Рекомендуемая литература по дисциплине «Молекулярная медицина»:
1.Северин Е.С., Биохимия [Электронный ресурс]: учебник / под ред. Е. С. Северина. - 5-е изд., испр. и доп. - М. : ГЭОТАР-Медиа, 2016. - 768 с. - ISBN 978-5-9704-3762-9 - Режим
доступа: http://www.studmedlib.ru/book/ISBN9785970437629.html
2.Авдеева Л.В., Биохимия [Электронный ресурс] / Авдеева Л.В., Алейникова Т.Л., Андрианова Л.Е., Белушкина Н.Н., Волкова Н.П., Воробьева С.А., Голенченко В.А., Губарева А.Е., Корлякова О.В., Лихачева Н.В., Павлова Н.А., Рубцова Г.В., Силаева С.А., Силуянова С.Н., Титова Т.А. - М. : ГЭОТАР-
Медиа, 2014. - 752 с. - ISBN 978-5-9704-3043-9 - Режим доступа: http://www.studmedlib.ru/book/ISBN9785970430439.html
3.Губарева А.Е., Биологическая химия. Ситуационные задачи и тесты [Электронный ресурс] : учеб. пособие / А. Е. Губарева [и др.] ; под ред. А. Е.
Губаревой. - М. : ГЭОТАР-Медиа, 2016. - 528 с. - ISBN 978-5-9704-3561-8 -
Режим доступа: http://www.studmedlib.ru/book/ISBN9785970435618.html
Тема: «Синтез глюкозы из веществ неуглеводной природы -
глюконеогенез, регуляция»
Цель занятия: изучить синтез глюкозы в процессе глюконеогенеза,
определить его связь с другими обменами.
План:
1.Глюконеогенез: характеристика процесса, субстраты,
локализация процесса, биологическое значение.
2.Необратимые реакции гликолиза и глюконеогенеза. Схема глюконеогенеза в организме человека, этапы, ферменты.
3.Синтез глюкозы из лактата и глицерина, аминокислот. Циклы:
Кори и глюкозо-аланиновый.
4.Регуляция процессов гликолиза и глюконеогенеза.
1. Глюконеогенез: характеристика процесса, субстраты,
локализация процесса, биологическое значение.
Глюконеогенез – процесс синтеза глюкозы «de novo» из веществ неуглеводной природы.
Некоторые ткани, например мозг, нуждаются в постоянном поступлении глюкозы. Когда поступление углеводов в составе пищи недостаточно,
содержание глюкозы в крови некоторое время поддерживается в пределах нормы за счёт расщепления гликогена в печени. Однако запасы гликогена в печени невелики. Они значительно уменьшаются к 6-10 ч голодания и практически полностью исчерпываются после суточного голодания. В этом случае в печени начинается синтез глюкозы de novo
Биологическая роль глюконеогенеза - поддержание уровня глюкозы в крови в период длительного голодания и интенсивных физических нагрузок.
Локализация: главным образом в печени и менее интенсивно - в
корковом веществе почек, а также в слизистой оболочке кишечника. Эти ткани могут обеспечивать синтез 80-100 г глюкозы в сутки. На долю мозга при голодании приходится большая часть потребности организма в глюкозе. Это
объясняется тем, что клетки мозга не способны, в отличие от других тканей,
обеспечивать потребности в энергии за счёт окисления жирных кислот.
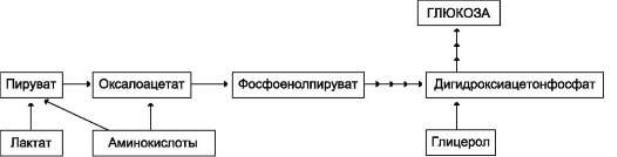
Первичные субстраты глюконеогенеза: лактат, аминокислоты и глицерол.
Включение этих субстратов в глюконеогенез зависит от физиологического состояния организма.
Лактат - образуется при любых состояниях организма в эритроцитах и работающих мышцах. Таким образом, лактат используется в глюконеогенезе постоянно.
Аминокислоты - образуются в результате распада белков мышц и соединительной ткани и включаются в глюконеогенез при длительном голодании или продолжительной мышечной нагрузке.
Глицерол - высвобождается при гидролизе жиров в жировой ткани в постабсорбтивный период или при физической нагрузке;
Рис.1 Включение субстратов в глюконеогенез: связь трех видов обмена веществ
2. Неообратимые реакции гликолиза и глюконеогенеза. Схема
глюконеогенеза в организме человека, этапы, ферменты.
Большинство реакций гликолиза и глюконеогенеза являются обратимыми и катализируются одними и теми же ферментами. Три реакции гликолиза (1,3 и 10-я) являются необратимыми, поэтому существуют обходные стадии глюконеогенеза и гликолиза. На этих стадиях реакции глюконеогенеза протекают другими путями.
I-я обходная стадия: образование ФЕП из пирувата.
II-я обходная стадия: гидролиз фруктозо-1,6-бисфосфата
III-я обходная стадия: гидролиз глюкозо6-фосфата.
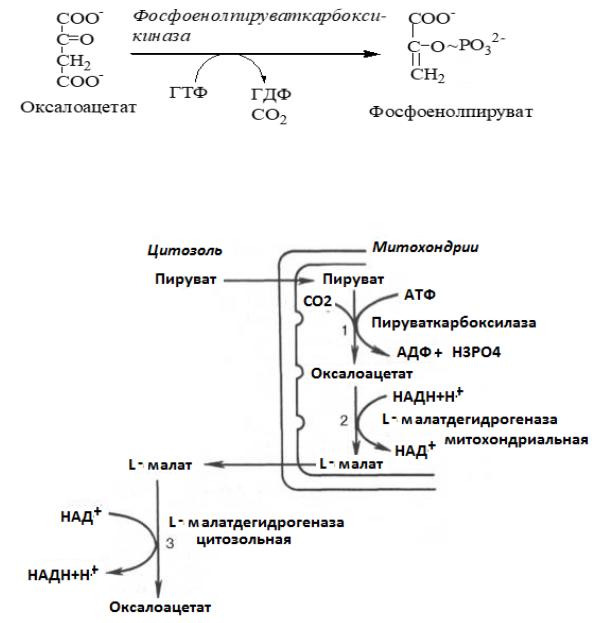
I-я обходная стадия: образование фосфоенолпирувата из пирувата состоит из 4-х реакций. В митохондриях пируваткарбоксилаза превращает ПВК в оксалоацетат. Эта реакция идет в клетке постоянно, являясь анаплеротической (пополняюшей) реакцией ЦТК.
СОО |
- |
Пируваткарбоксилаза |
|
- |
||
|
|
|
|
|||
|
СО |
, биотин |
|
СОО |
||
|
|
2 |
|
|
С O |
|
С O |
|
|
|
|
|
|
|
|
|
|
СН |
|
|
|
|
|
|
|
|
|
СН |
|
|
|
|
2 |
- |
3 |
|
АТФ |
АДФ, Н РО |
|
СОО |
|
|
|
4 |
||||
Пируват |
|
3 |
Оксалоацетат |
|||
|
|
|
||||
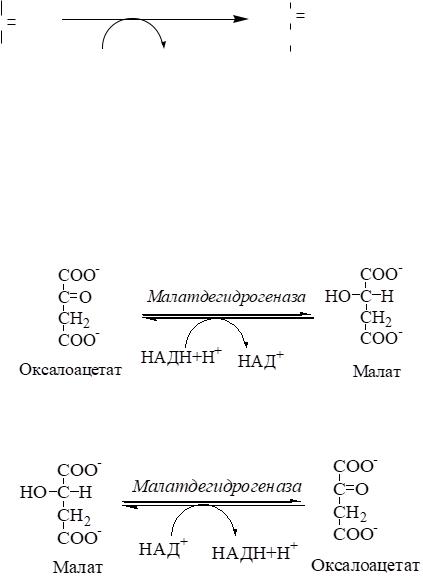
Мембрана митохондрий непроницаема для оксалоацетата.
оксалоацетата. Зато через мембрану может пройти малат. Повернуть малатдегидрогеназную реакцию ЦТК вспять позволяет избыток НАДН в митохондриях. НАДН поступает из β-окисления жирных кислот,
активируемого в условиях недостаточности глюкозы в гепатоците.
В результате малат накапливается, выходит в цитозоль и здесь превращается обратно в оксалоацетат.
В цитоплазме фосфоенолпируваткарбоксикиназа осуществляет превращение ОАА в ФЕП, для реакции требуется энергия ГТФ. От молекулы отщепляется тот же углерод, что и присоединяется.
После образования фосфоенолпирувата все остальные реакции также протекают в цитозоле вплоть до образования фруктозо-1,6-бисфосфата и катализируются гликолитическими ферментами.
Рис.2 Схема превращения пирувата в ФЕП.
II-я обходная стадия: гидролиз фруктозо-1,6-бисфосфата
Второе препятствие на пути синтеза глюкозы – фосфофруктокиназная реакция – преодолевается с помощью фермента фруктозо-1,6-дифосфатазы.
Этот фермент есть в почках, печени, поперечно-полосатых мышцах. Таким образом, эти ткани способны синтезировать фруктозо-6-фосфат и глюкозо-6-
фосфат.
III-я обходная стадия: гидролиз глюкозо-6-фосфата.
Последняя реакция катализируется глюкозо-6-фосфатазой. Она имеется только в печени и почках, следовательно, только эти ткани могут продуцировать свободную глюкозу.
3.Синтез глюкозы из лактата и глицерина, аминокислот. Циклы:
Кори и глюкозо-аланиновый.
Синтез глюкозы из пирувата. В ходе этого процесса расходуются 6
моль АТФ на синтез 1 моль глюкозы из 2 моль пирувата. Четыре моль АТФ расходуются на стадии синтеза фосфоенолпирувата из оксалоацетата и ещё 2
моль АТФ на стадиях образования 1,3-бисфосфоглицерата из 3-фосфо-
глицерата.
Суммарный результат глюконеогенеза из пирувата выражается следующим уравнением:
2 Пируват + 4 АТФ + 2 ГТФ + 2 (NАDН+ Н+) + 4 H2O →
Глюкоза + 4 АДФ + 2 ГДФ + 6 Н3РО4 + 2 NАD+.
Синтез глюкозы из аминокислот. В условиях голодания часть белков мышечной ткани распадается до аминокислот, которые далее включаются в процесс катаболизма.
Аминокислоты, которые при катаболизме превращаются в пируват или метаболиты цитратного цикла, могут рассматриваться как потенциальные предшественники глюкозы и гликогена и носят название гликогенных.
Например, оксалоацетат, образующийся из аспарагиновой кислоты, является промежуточным продуктом как цитратного цикла, так и глюконеогенеза.
Из всех аминокислот, поступающих в печень, примерно 30% приходится на долю аланина.
Это объясняется тем, что при расщеплении мышечных белков образуются аминокислоты, многие из которых превращаются сразу в пируват или сначала в оксалоацетат, а затем в пируват. Последний превращается в аланин, приобретая аминогруппу от других аминокислот.
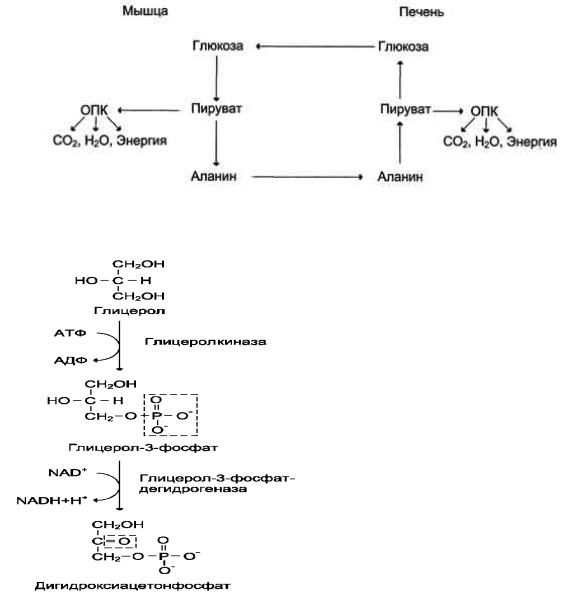
Аланин из мышц переносится кровью в печень, где снова преобразуется в пируват, который частично окисляется и частично включается в глюконеогенез.
Следовательно, существует следующая последовательность событий
(глюкозо-аланиновый цикл): глюкоза в мышцах → пируват в мышцах → аланин в мышцах → аланин в печени → глюкоза в печени → глюкоза в мышцах.
Весь цикл не приводит к увеличению количества глюкозы в мышцах, но он решает проблемы транспорта аминного азота из мышц в печеньи предотвращает лактоацидоз.
Рис.3 Цикл аланин-глюкоза
Синтез глюкозы из глицерола.
Рис.4 Образование дигидроксиацетонфосфата
Глицерол образуется при гидролизе триацилглицеролов,
главным образом в жировой ткани. Использовать его могут только те ткани, в которых имеется фермент глицеролкиназа, например печень, почки. Этот АТФ-
зависимый фермент катализирует превращение глицерола в α-глицерофосфат
(глицерол-3-фосфат).