

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfвстала проблема снабжения пищей слабоосвещенных, находящихся на большой глубине час тей, где фотосинтез отсутствовал. Появились специализированные проводящие питательные вещества ткани, пронизывающие таллом, связы вающие его верхние части с нижними нефотосинтезирующими.
Следовательно, характерные признаки много- клеточности—качественная неравноценность кле ток, слагающих тело, их дифференцировка и объ единение в комплексы различной сложности (ткани, а позднее органы), выполняющие разные функции в целостном организме. Для многокле точных характерно также индивидуальное разви тие (онтогенез), начинающееся в большинстве случаев, исключая вегетативное размножение, с деления одной клетки (зиготы, споры). Тенден ции к переходу в многоклеточное состояние воз никали в процессе эволюции у водорослей, бак терий, простейших (многоядерность, колониальность и т. д.), но остались до конца незавер шенными.
Существуют различные гипотезы происхожде ния многоклеточности, например теория закре пившегося симбиоза одноклеточных, теория фагоцителлы и ряд других, однако единого мнения по этому вопросу нет. Наиболее вероятно, что многоклеточные организмы возникли впервые в результате усложнения организации некоторых колониальных простейших.
Простейшей формой многоклеточного слое вища у водорослей является нитчатая структура. Она представлена талломами, состоящими из не скольких или очень многих клеток, расположен ных в виде нити. Нити могут быть простыми и разнообразно ветвящимися, свободноживущими, прикрепленными и объединенными в слизистые колонии (рис. 308, 1—6).
Клетки в нитчатых слоевищах тесно связаны друг с другом, во многих случаях здесь обнаруже ны поры и плазмодесмы на поперечных стенках. В то же время нити могут распадаться на участки
и даже на отдельные клетки, что является обыч ным способом вегетативного размножения мно гих нитчатых водорослей.
У более просто организованных нитчатых во дорослей талломы слагаются из одного ряда по добных друг другу клеток (рис. 308, I). Наряду с этим у многих водорослей нити изменяются к концам — утончаются (рис. 308, 3), расширяются или заканчиваются своеобразной по форме клет кой. Наиболее отчетливо различия в строении нитей выражены при прикрепленном образе жиз ни, определяющем полярность таллома. Нередко нижняя клетка превращается в ризоид, или стопу (рис. 308, 2).
По-разному происходит рост нитчатого слое вища. Если все клетки таллома делятся, рост на зывают диффузным (см. рис. 308, 1). В других слу чаях рост нити происходит только в определен ных, меристемальных, зонах. В зависимости от ее расположения различают интеркалярный рост — зона роста расположена в средней части нити;
апикальный, или верхушечный, и базальный — рост основанием (рис. 308, 4—6).
Нитчатая структура в эволюционном отноше нии интересна как исходная ступень для образо вания более сложных многоклеточных структур, присущих многим водорослям.
Некоторые особенности нитчатого таллома (полярность, дифференцировка клеток, типы рос та) закрепились в процессе эволюции у более вы сокоорганизованных водных растений, а впослед ствии и у наземных.
Дальнейшим эволюционным шагом диффе ренциации тела водорослей является разнонитчатая структура. Талломы данной структуры, яв ляясь нитчатыми в своей основе, подразделяются на две части: стелющуюся по субстрату, горизон тальную, и прямостоящую, вертикальную (рис. 309).
Разнонитчатая структура представлена в отде лах сине-зеленых, зеленых, золотистых, бурых и красных водорослей, где она является или посто янной формой существования, или временным состоянием. Талломы этой структуры у красных и бурых водорослей имеют многометровые размеры и довольно высокую специализацию.
Рис. 308. Нитчатая структура у сине-зеленых водорослей: |
|
|
|
/ — простейшее строение нити с диффузным ростом у Oscillatoria; 2 — нить с |
|
||
дифференцированным основанием у Endonema; 3 — верхушка нити у |
|
||
Rivularia, вытянутая в волосок; 4 — интеркалярный рост у Gloeotrichia; 5 — |
Рис. 309. Разнонитчатая структура у сине-зеленой водоросли |
||
апикальный рост на концах ветвей у Scytonema; 6 — базальный |
рост у |
||
Fischerella |
|||
Calothrix |
|
||
342

Рис. 310. Пластинчатая структура у зеленой водоросли Prasiola: однорядная нить, разрастающаяся в однослойную пластинку
Интересна с точки зрения эволюции пластин чатая структура таллома. Она характеризуется многоклеточными слоевищами в форме пласти нок, состоящих из одного, двух или нескольких слоев клеток (рис. 310). Образование их всегда начинается с нити и происходит в результате продольного деления клеток.
Пластинчатые водоросли растут либо свобод но распростертыми по субстрату, либо прикреп ленными к нему в одном месте края пластинки. Они хорошо представлены в отделах зеленых, бу рых и красных водорослей, достигая порой зна чительных размеров.
Таким образом, рассматривая эволюционный путь дифференциации тела у водорослей, нельзя не заметить, что мир водорослей огромен и зани мает в растительном царстве особое, исключи тельное по своему значению место как в истори ческом аспекте, так и по той роли, которая при надлежит ему во всем последующем развитии растений и в общем круговороте веществ в при роде. В общем плане водоросли характеризуются талломным (слоевцовым) строением, наличием хлорофилла, размножением с помощью спор (или вегетативно), т. е. это низшие споровые растения.
Возникновение хлорофилла у первичных бес цветных гетеротрофных организмов было на столько существенным переломным моментом, что только его и можно считать началом той бо гатейшей эволюции, которая привела к созданию биосферы на Земле в ее современном выражении. И трудно допустить, что такое сложное и уни кальное органическое соединение, как хлоро филл, давшее организмам возможность фототрофного питания, т. е. совершенно новой энер гетики жизненных процессов, основанной на утилизации солнечной энергии, могло возник нуть многократно на разных этапах эволюции. С этих позиций мир водорослей един и уникален. Морфологическое разнообразие его различных рассмотренных выше ветвей есть следствие эво люционного взрыва, вызванного появлением фо тосинтеза, который обеспечил хлорофиллонос-
ным организмам успешное развитие в чисто абиотической среде.
В процессе эволюционного развития в группе водорослей появились и закрепились многие спе цифические черты структурно-функциональной организации, которые присущи типичным пред ставителям Царства растений, а именно:
1)фототрофный способ питания;
2)наличие упругой полисахаридной оболочки, образующей опорную систему, наружный скелет клетки;
3)специфические черты организации клетки;
4)дифференциация тела, появление многоклеточности;
5)морфоструктурное разнообразие организа ции тела (нитчатые, разветвленные, уплощенные талломы, ризоиды, полярность);
6)появление специализированных групп кле ток (тканей);
7)способность к длительному нарастанию за счет образовательных клеток (верхушечный, интеркалярный, базальный рост), т. е. открытая сис тема роста;
8)различные типы ветвления талломов: а) виль чатое, при котором оси подчиненного порядка таллома закладываются на верхушке материнской оси, клетка которой может дать начало сразу двум— дихотомия, трем — трихотомия или нескольким (политомия) осям; б) боковое, когда на главной оси, несколько отступив от ее окончания, возни кают боковые оси, каждая из которых способна образовывать свои боковые оси. При этом спосо бе ветвления потенциальные возможности фор мирования новых систем таллома более разнооб разны и биологически более выгодны. Возникно вение и развитие боковых ответвлений может происходить в направлении от основания к вер хушке оси (акропетально) либо от верхушки ор гана к его основанию (базипетально). Описанные типы ветвления, видимо, возникали многократно
инезависимо друг от друга в различных линиях эволюции низших растений;
9)вегетативное, бесполое (спорами) и половое (гологамия, изогамия, анизогамия, оогамия) раз множение. Органы размножения одноклеточные, но у харовых уже многоклеточные;
10)жизненный цикл водорослей включает че редование спорофита и гаметофита. Наблюдаются самые различные взаимоотношения диплоидной
игаплоидной фаз жизненного цикла1.
Обобщая изложенное, можно утверждать, что водоросли — одни из древнейших организмов, населяющих нашу планету. Пожалуй, только бак терии могут поспорить с ними в древности про исхождения и длительности существования. В прошлые геологические эпохи, как и в настоящее время, водоросли населяли океаны, реки, озера и другие водоемы. Обогатив атмосферу кислородом, они вызвали к жизни разнообразный мир живот ных и явились родоначальниками растений, засе ливших сушу. Рис.311 наглядно демонстрирует
1 В обзоре не рассматривается эволюция размножения у водорослей, так как этот сложный и специфический вопрос требует специального изучения.
343

эволюционный путь их развития (Ю. Е. Пет ров И Жизнь растений. М., 1977. Т. 3. С. 353).
Следует отметить, что вопрос происхождения наземных растений сложный и до конца не вы ясненный. Поэтому существует целый ряд ги потез, в которых наземные формы выводят из разных групп водорослей—зеленых, бурых, харовых и т. д.
ВЫХОД РАСТЕНИЙ НА СУШУ, ВОЗНИКНОВЕНИЕ ВЕГЕТАТИВНЫХ ОРГАНОВ, ТКАНЕЙ
Эволюционные возможности первичных водо емов (океанов, морей и т. д.) не могли сохранять ся вечно. Типичные водные растения — водорос ли — прекрасно приспособлены к среде обита ния. В процессе эволюции они выработали высокоорганизованные признаки внешней и внутренней структуры, питания, размножения, позволяющие в совершенстве использовать усло вия водной среды. Но в процессе эволюции мор ских организмов между ними возникала все более сильная конкуренция за место под солнцем (среди животных за пищу и за убежища). Пред ставители многих групп водорослей (а также бак терий, грибов) заселяли реки, озера, однако об щая площадь этих пресноводных местообитаний очень мала, а кроме того, они время от времени пересыхали. Рядом с морем была суша, по выра жению К. Даддингтона (1972), растениям «необ ходимо было ее завоевать». Суша в те времена — это бесплодные скалы и пустыни, на которых от сутствовала почва, если не считать рассеянной кое-где пыли. Но и лишенная почвы суша пред ставляла благоприятные возможности для жизни растений: здесь было изобилие света, кислорода и углекислого газа, которые циркулируют в воздухе быстрее, чем в воде; горные породы, образован ные из тех же минеральных веществ, которые не обходимы растениям. На суше для растений вна чале не было конкуренции.
Однако на суше существовали проблемы, изза которых она долгое время оставалась необи таемой. Самой главной проблемой было высыха ние, так как в воздушной среде вода легко испа ряется. Таким образом, вода явилась критиче ским фактором при освоении суши живыми организмами.
Другое осложнение, сопряженное с жизнью на суше, обусловлено тем, что ввиду разной плотно сти воздуха и воды гидростатическая сила, помо гающая уравновесить массу организма, в воздухе гораздо меньше, чем в воде. Поэтому организмам, находящимся в воздушной среде, необходимы прочные опоры (скелеты), иначе они никогда не «оторвутся» от Земли.
Почему-то заселение суши растениями далеко не всеми исследователями признавалось пробле мой. Некоторым казалось, что не над чем ломать голову — растения вышли из моря на сушу, сна чала заселив мелководья, а потом двинулись вдоль рек в глубь континентов. Но накопленные палеоботанические данные показывают, что все было не так просто (С. В. Мейен, 1989; В. А. Красилов, 1989).
Считается, что первые растения появились на суше в силурийский период, т. е. более 300 млн лет назад. Однако в отложениях силура найдены остатки высших наземных растений. С. В. Мейен (1989), обобщив работы многих современных па леоботаников, считает, что на суше сначала посе лились не высшие растения, а различные водо росли, грибы, бактерии, образовав вместе назем ные экосистемы. Так, американский палеобота ник Г. Реталляк в палеопочве позднего ордовика (второй период палеозоя, следует за кембрий ским, предшествует силурийскому периоду, 490— 435 млн лет назад) в штате Пенсильвания обна ружил ходы животных, уходящие в глубь почвы с поверхности. Скорее всего это были какие-то кольчатые черви. Но раз они жили на суше, зна чит чем-то питались. Приходится допустить, что растительный покров, поддерживающий назем ную фауну, существовал и до силура, т. е. в ордо вике. Г. Реталляк считает, что это были какие-то несосудистые растения — водоросли, поскольку в палеопочве нет их подземных частей. Выходит, ордовикская суша была заселена водорослевой наземной флорой, несомненно, продуктивной, так как могла прокормить многоклеточных — до вольно крупных животных. С. В. Мейен (а также Г. Стеббинс, Г. Хилл) высказывает предположе ние о происхождении высших растений от назем ных водорослей. «Корни» этой предполагаемой на земной несосудистой растительности уходят в конец архея — начало протерозоя: уже с раннего докембрия известны палеопочвы, а в одной из них (возраст около 2,4 млрд лет) обнаружено ор ганическое вещество. В докембрии приливноотливная зона была заселена микроорганизмами, часть которых похожа на современные формы, живущие в наземных водорослевых корках. В. Райт считает, что в раннем палеозое уже могли существовать лишайники. Аскомицеты, т. е. те грибы, которые в большинстве случаев образуют симбиотические сообщества с водорослями у со-
344
временных лишайников, известны по крайней мере с силура.
Этот небольшой экскурс в современную па леоботанику показывает, насколько сложна про блема заселения суши растениями. Пока общей точки зрения у специалистов нет. Но гипотеза о том, что первыми сушу освоили низшие расте ния, в частности водоросли, приобретает все больше сторонников.
Так или иначе, первыми завоевателями суши были, вероятно, растения (и животные), жившие в приливно-отливной зоне или в периодически пересыхающих водоемах, т. е. организмы, уже приспособившиеся к коротким периодам сущест вования вне воды. Их привыкание к суше шло очень медленно, они постепенно передвигались к берегу, пока не оказались выше границы прили ва, где на них попадали только брызги — в силь ный шторм или при исключительно высоком уровне воды весной. В настоящее время такую же картину можно наблюдать у водоросли Pelvetia canaliculata, которая растет так высоко на берегу, что является почти сухопутным растением. Отсю да был только один маленький шаг до полного отрыва от моря, но включал он в себя серьезную перестройку всей структуры и образа жизни рас тений, выходящих на сушу, и явился поворотным событием в морфологической эволюции расти тельного мира.
Приспособление крупных многоклеточных форм к обитанию в почвенно-воздушной среде привело к появлению высших растений. Различия между низшими растениями, до этого уже засе лившими сушу, и большинством высших расте ний заключаются в том, что низшие растения обитают в среде, насыщенной парами воды, а при наступлении засухи теряют воду, пересыхают и впадают в анабиоз (греч. anabiosis — оживление, возвращение к жизни). У низших наземных рас тений водный обмен не стабилизирован и интен сивность их жизненных процессов полностью за висит от присутствия влаги в окружающей среде.
Высшие наземные растения стабилизировали содержание воды внутри своего тела и стали от носительно независимыми от колебания влажно сти в атмосфере и почве.
Наземные условия существования резко отли чаются от условий жизни в воде. На земле расте ние живет одновременно в двух существенно раз ных средах, поэтому его тело оказалось разделен ным на две части — подземную и надземную, вьшолняющие различные функции. Подземная часть обеспечила почвенное питание (всасывание воды и растворенных в ней веществ), но потеряла способность к фотосинтезу, тогда как надземная часть растения осуществляла автотрофный способ питания (фотосинтез), но утратила непосредст венную связь с влагой почвы. Разделение функ ций обусловило появление специализированных групп клеток — тканей, восполняющих многие из тех функций, которые в водной среде выполняет вода. Таким образом, органы и ткани появились в группе наземных высших растений в связи с их приспособлением к новым условиям обитания после выхода на сушу.
РАННИЕ ЭТАПЫ МОРФОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ НАЗЕМНОЙ ФЛОРЫ
Морфологическая эволюция первых высших растений Земли довольно достоверно изучена на основе данных палеоботаники.
Интересные свидетельства о первичном расти тельном населении суши дают остатки спор, ко торые извлекают как из прибрежных отложений морей, так и из континентальных отложений.
Анализ структуры спор. позволил палеобота никам выделить три главных этапа развития древней наземной растительности. Первый начи нается в середине ордовика, когда споровые рас тения впервые появились на суше. Эти древние споры сохранились в тетрадах, обтянутых общей оболочкой, и напоминают тетрады спор печеноч ников в мхов. Однообразие и обилие спор указы вают на незначительное разнообразие материн ских растений и в то же время на быстрое освое ние ими суши. На втором этапе, в середине или верхней части нижнесилурийских отложений, однообразные тетрады сменяются одиночными спорами, которые напоминают споры высших споровых растений. В начале третьего этапа (се редина верхнего силура) преимущественно гладкостенные споры сменяются спорами с разнооб разным рисунком оболочки. Эти три этапа про слеживаются на достаточно большой площади —
вАнглии, Африке, Северной и Южной Америке.
В1859 г. канадский геолог Дж. Досон в де вонских отложениях на п-ве Гаспе в Канаде об наружил остатки примитивного высшего расте ния, не похожего ни на одно из известных науке
вто время. Растение имело дихотомически раз ветвленные стебли, лишенные листьев, проводя щую систему в виде протостелы. Спорангии этого растения были верхушечными: они свисали с верхушек повторно дихотомированных боковых веточек. Подземные органы, к сожалению, обна ружены не были. Дж. Досон дал растению назва ние Psilophyton princeps (псилофит), что можно перевести как «голорос первичный» (рис. 312, 7). К сожалению, находка Дж. Досона осталась неза меченной, и ее значение для познания эволюции высших растений не было оценено. Слишком отличался псилофит от известных растений и не укладывался в существовавшие тогда системы классификации. Положение изменилось только в 20-е годы нашего века.
В1912 г. близ деревни Райни (Шотландия) врач и геолог-любитель У. Макки открыл пласты кремнистой породы чёрта (англ. chert), известно го в литературе под названием «райниевого чёрта», а в них хорошо сохранившиеся окремнелые растительные остатки. У. Макки передал для изучения все материалы выдающемуся шотланд скому палеоботанику Р. Кидстону (1852—1924), который совместно с профессором Манчестер ского университета У. Лангом в 1917—1921 гг. опубликовал пять работ, вошедших в историю ботаники как подлинно классические исследова-
23 Зак. 1851 |
345 |

Рис. 312. Первичная наземная флора:
1 — псилофит первичный (Psilophyton princeps); 2 — риния большая (Rhynia major): a — внешний вид (реконструкция); б — попереч ный срез стебля (в центре стебля видна протостела); 3 — куксония (Cooksonia); 4 — зостерофиллум (Zosterophyllum); 5 — госслингия (Gosslingia)
ния, положившие начало систематическому изу чению древнейших и наиболее примитивных высших растений.
Окремнелые остатки одних из наиболее древ них представителей высших растений, сохранив шихся в пластах «райниевого чёрта» в раннем де воне (не ранее 415 млн лет назад), благодаря спе цифическим условиям сохранились на месте своего произрастания, в вертикальном положе нии, хорошо демонстрируя внешнюю форму и тончайшие детали внутреннего строения.
Р. Кидстон и У. Ланг установили ряд родов (риния — Rhynia, хорнеофит — Horneophyton и др.), имевших определенное сходство с псилофитом (рис. 312, 2, а, б).
В 1937 г. У. Ланг в верхнесилурийских песча никах Уэльса (Великобритания) открьш новый род, названный им куксонией (рис. 312, 3). Позднее куксония была обнаружена в верхнеси лурийских отложениях Чехии, США, а также в нижнем девоне Шотландии и Западной Сибири. Куксония — самая древняя из достоверных нахо док высших растений. Это растение жило более 415 млн лет назад. Его и помещают обычно в ос нование филогенетического древа всех высших растений.
В результате исследований, начатых Дж. Досоном, Р. Кидстоном и У. Лангом, оказалось, что
риния, хорнеофит, куксония, псилофит и рад близких родов (тениокрада, хедея, яравия) со ставляют наиболее древний и самый примитив ный отдел высших растений, геологическая исто рия которого начинается с силурийского периода и заканчивается в позднем девоне. Ранее этот от дел обозначали псилофитовые (Psilophyta), но в настоящее время многие ученые предпочитают название риниофиты (Rhyniophyta).
Риниофиты представляли собой небольшие травянистые растения. Например, у ринии, наи более полно и детально изученного представителя риниофитов, цилиндрические органы ветвились дихотомически и имели апикальное нарастание. Хорошо изучен у этих растений горизонтальный корневищеобразный орган (А. Л. Тахтаджян на звал его «ризомоид»), от которого вверх отходили стебли, а вниз — многочисленные ризоиды, всасы вающие воду с растворенными в ней минераль ными веществами. В общем риниофиты характе ризовались верхушечным расположением споран гиев, отсутствием корней и листьев, дихотомиче ским или псевдомоноподиальным ветвлением спорофита, очень примитивным анатомическим строением и разноспоровостью. Проводящая сис тема была развита слабо и представляла типич ную протостелу. Ксилема состояла из трахеид с кольчатым, спиральным или реже лестничным (у
346
псилофита) утолщением. В отличие от большин ства сосудистых растений механические (опор ные) ткани отсутствовали. Имелись немногочис ленные и просто устроенные устьица, лишенные сопровождающих клеток в эпидермисе (кожице), покрывающем стебель. Под эпидермисом залега ла хлорофиллоносная ткань, способная к фото синтезу. Риниофиты не обладали еще способно стью ко вторичному росту (меристема у них была только апикальная). Спорангии были толстостен ные, разные по форме и размерам, у ринии они еще слабо отличались от верхушек обычных сте рильных веточек. Более того, у хорнеофита они были однажды или дважды вильчатые.
В спорангиях риниофитов развивались много численные споры, по строению оболочки типич ные для высших растений. Вопрос о гаметофите риниофитов пока не ясен.
Есть основания предполагать, что жили они в теплом или даже жарком климате с сезонными дождями, вероятно, непосредственно у уреза во ды, образуя одновидовые заросли по берегам крупных, часто разливавшихся рек, а также в прибрежном мелководье. Некоторые из них, на пример тенпокрада, были водными растениями с поднимающимися над поверхностью воды споро носными частями. Но были среди них, несо мненно, и вполне сухопутные представители.
Имеются все основания считать риниофиты той исходной предковой группой, от которой произошли моховидные, хвощевидные и папо ротниковидные, а среднедевонские растения из родов зостерофиллум и госслингия отдела зостерофиллофиты (Zosterophyllophyta), близкого к отделу риниофитов, дали, вероятно, начало отде лу плауновидных (рис. 312, 4, 5).
Таким образом, проанализировав этот этап эволюции растений, можно предположительно утверждать, что высшие растения произошли от каких-то водорослей. Об этом говорит прежде всего то, что в геологической истории раститель ного мира эре высших растений предшествовала эра водорослей. В пользу данного предположения говорят также следующие факты: 1) сходство наиболее древней и уже давно вымершей группы высших растений — риниофитов — с водоросля ми и, в частности, очень сходный характер их ветвления; 2) сходство в чередовании поколений высших растений и многих водорослей; 3) нали чие жгутиков и способность к самостоятельному плаванию у мужских половых клеток многих высших растений; 4) сходство в строении и функциях хлоропластов. Некоторые ученые вы сказали предположение, что переходу водоросле вого предка высших растений в условия назе*шого существования способствовал симбиоз с гри бами.
ЭВОЛЮЦИЯ ВЫСШИХ РАСТЕНИЙ
Эволюционное развитие высших растений де тально изучается в курсе систематики. Однако основные направления морфологической эволю
ции растений можно объяснить только с точки зрения общей линии эволюции, которую кратко можно представить следующим образом.
Резкая дифференциация растительных форм произошла в среднем девоне, что привело к воз никновению существовавших в дальнейшем ос новных групп папоротниковидных. На смену низкорослым риниофитам (1—3 м) пришли дре вовидные высокорослые формы плаунов, хвощей, достигающие в высоту 40 м. В познедевонскую эпоху и в карбоне (каменноугольном периоде) в течение относительно короткого периода появи лись представители многих таксономических групп папоротниковидных, которые стали господ ствовать на пригодной для расселения растений части суши. Земля начала зеленеть. Это время по праву называют временем папоротниковидных. Остатки древовидных растений, образовывавших настоящие леса, обнаружены уже в отложениях позднедевонской эпохи. Наиболее пышного раз вития растительный покров достиг в позднекаменноугольную эпоху. Высокие древовидные плауны, хвощи (некоторые из них размножались семенами), папоротники и семенные папоротни ки образовывали пышные растительные сообще ства, напоминавшие растительный мир совре менных тропиков. В это же время появились хвойные и другие голосеменные растения.
Несколько позже, в пермский период, почти повсюду на суше стали господствовать представи тели разных таксономических групп голосеменных растений, которые вытеснили доминировавших до тех пор представителей папоротниковидных; началось время голосеменных. Эта резкая смена флор, как полагают, в значительной мере связана с изменением климата. Климат стал суше, и голо семенные, которые имели более мощную систему тканей, проводящих воду, оказались лучше при способленными к наземным условиям. На протя жении всей мезозойской эры, вплоть до раннемеловой эпохи, характер растительности определяли представители разных групп голосеменных: са говниковые, гинкговые, кейтониевые, беннеттитовые, а также хвойные. В настоящее время со хранились лишь немногочисленные остатки этого многообразия; в мезозойскую эру многие из них были распространены почти повсеместно. Сейчас саговниковые насчитывают всего около 90 видов, гинкговые — только один, а из 40 000 видов беннеттитовых не осталось ни одного.
ОСНОВНЫЕ ОРГАНЫ ВЫСШЕГО РАСТЕНИЯ
Изучение строения риниофитов и их эволю ционных взаимоотношений имеет исключитель ное значение для эволюционной морфологии и филогении всех высших растений. Оно прежде всего показало, что первоначальным органом высших растений был телом (греч. telos — ко нец), являющийся спорофитом. Он представлял собой дихотомически ветвящийся стебель, несу щий верхушечные спорангии, в то время как корни и листья отсутствовали и произошли, сле-
347

довательно, позже спорангия и стебля. В ходе дальнейшего приспособления к наземной жизни риниофиты дали начало растениям, обладавшим корнями и побегами. Риниофиты представляют основной объект теломной теории происхождения и эволюции спорофита высших растений. Со гласно теломной теории, созданной В. Циммер маном (1930, 1965), которой придерживаются многие современные ботаники, высшие растения с настоящими корнями и побегами (стеблями с расположенными на них листьями и почками) возникли из совокупности теломов риниофитов в результате перевершинивания, уплощения, сра стания и редукции теломов. Листья папоротников (вайи) и семенных растений возникли из упло щенных и сросшихся между собой систем тело мов («плосковетки»). На рис. 313 показано, как,
Рис. 313. Схема возникновения плоского листа из группы те ломов:
1—4 — последовательные этапы, от более примитивных к более продвинутым группам растений (по А Е. Васильеву и др., 1978)
по современным представлениям, основанным на изучении ископаемых остатков вымерших расте ний, плоские ассимилирующие листья в процессе эволюции возникли из групп теломов путем их
расположения в одной плоскости, скучивания и сра стания. Отражение этого процесса можно видеть в жилковании листьев у многих ископаемых и современных папоротников (рис. 314).
Стебли с их сложной проводящей системой, вероятно, возникли в результате бокового сраста ния теломов, корни — из систем подземных те ломов (ризомоидов).
Таким образом, листостебельный побег обра зовался в результате объединения групп теломов и постепенной дифференциации их в связи с различными функциями на цилиндрические осе
лке. 314. Сегмент листа папоротника Адиантум венерин волос (Adiantura capillus-veneris) с дихотомическим жилкованием
вые органы (стебли) и плоские боковые органы (листья). В ходе эволюции стебель сохранил спо собность к длительному верхушечному нараста нию и ветвлению, а листья эту способность утра тили (исключение — вайи папоротников).
Успешной колонизации суши высшими рас тениями способствовало возникновение почек - замкнутых влажных вместилищ, защищенных эм бриональными листьями и почечными чешуями, где «работает* апикальная меристема. Благодаря такой защищенности меристем побеги смогли нормально функционировать (нарастать, ветвить ся) в условиях резких колебаний температуры и влажности.
Эволюция корня как осевого цилиндрическо го органа связана с развитием громадной всасы вающей поверхности путем многократного ветв ления и формирования систем корня.
Спороносные теломы эволюционировали не зависимо от побегов и корней. Таким образом, в результате усложнения тела растений при выходе на сушу и освоении воздушной и почвенной сред возникли основные органы растений — вегета тивные.
В современной ботанике под органами (греч. organon — орудие, инструмент) понимают части тела высших растений, которые имеют опреде ленное строение и выполняют определенную функцию.
Основными вегетативными органами растений являются побег (cormus) и корень (radix).
Основные органы тела высших растений (корень, стебель, лист) заложены уже в зародыше, поэтому классическая морфология выдвинула учение о трех органах тела — корне, стебле и лис те. Однако в настоящее время высказывается предположение, что высшие растения имеют два основных органа: корень и побег — совокупность стебля, листьев и почек. При этом стебель явля ется осевой (ортотропной), а листья — боковой (плагиотропной) частью побега.
Такой взгляд логичен при рассмотрении одно летних побегов. Но многолетние необлиственные скелетные оси деревьев и кустарников (стволы и стволики) трудно отождествить с побегами. В от ношении происхождения побега и первенства в его формировании стеблевой или листовой осно вы существуют три теории.
1. Главная часть побега — стебель; листья - производные стебля, его придатки. Эта теория основывается на структуре первых высших расте ний риниофитов. Как говорилось выше, надзем ные части этих растений были представлены без листными дихотомически ветвящимися осями - теломами. Не имели настоящих листьев и наибо лее примитивные виды плауновидных, производ ных риниофитов (ископаемые: содония украшен ная — Sawdonia ornata; каулангиофитон игли стый — Kaulangiophyton akantha и др.). Их стебли были покрыты мелкими боковыми выростами - шипами.
2. Главная часть побега — лист, а стебель формируется в результате срастания оснований листьев (черешков). Исходными группами расте ний, среди которых формировался такой исход-
348
ный побег, были древние папоротниковидные и саговниковые.
3. Среди частей побега нет преимущества. По бег состоит из многократно павторяющихся сход ных структур — узлов с междоузлиями, называе мых метамерами или фитонами.
В данной книге авторы придерживаются той точки зрения, что все элементы побега (стебель, листья и почки) онтогенетически возникают из единого комплекса меристемы и не функциони руют один без другого. Кроме того, побег как единое целое произошел от системы теломов и имеет единую проводящую систему. Поэтому по бег (а не его отдельные части — стебель, почки, листья) является универсальным надземным веге тативным органом.
Верно, однако, и представление о расчлене нии вполне сформированного побега на стебель (ось побега) и листья, характеризующиеся видоспецифической структурой и функцией. Эти час ти побега воспринимаются как его органы, т. е. как бы органы второго порядка.
И. Г. Серебряков (1952), К. И. Мейер (1958) предложили и обосновали схему расчленения те ла высшего растения, которая вошла в учебники (А. Е. Васильев и др., 1978, 1988).
- стебель
система побегов, ее элемент — побег ^___»_ листья
/ |
^*- почки |
i w v i a i n n i vv
_
х корневая система, ее элемент — корень
Система побегов вместе с корневой системой составляет тело плауновидных, хвощевидных, па поротниковидных и семенных растений. Возник новение побегов (т. е. листостебельности) — крупнейший ароморфоз (греч. airo — поднимаю и morphosis — образец, форма; эволюционное пре образование строения и функции организмов, имеющее общее значение для организма в целом и ведущее к морфологическому прогрессу) в ис тории растительного мира на Земле. Благодаря плоской форме листьев резко возросла фотосинтезирующая поверхность; связанное с этим уве личение транспирации способствовало развитию настоящих корней как совершенных органов по глощения воды и минеральных солей.
Побеги, несущие органы полового размноже ния, относятся к генеративным. Вместе с органа ми бесполого и вегетативного размножения они представляют репродуктивные органы (лат. ге — приставка, обозначающая повторение, возобнов ление действия и produco — произвожу).
ПРОБЛЕМЫ НАЗЕМНОЙ ЖИЗНИ. НАПРАВЛЕНИЯ МОРФОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Как только растения покинули водную среду обитания, они столкнулись с множеством про блем, требовавших разрешения. Эволюционная бо
таника, изучающая проблемы приспособления растений к сухопутным условиям, выделяет здесь целый ряд аспектов.
Прежде всего это проблема снабжения водой: для нормального функционирования раститель ный организм должен получать воду, без которой жизнь вообще невозможна. Для водорослей такой проблемы не существует, так как они чаще всего погружены в воду. Наземные животные, как пра вило, подвижны и могут отыскивать воду так же, как они отыскивают пищу. Грибы неподвижны, но живут в основном под поверхностью почвы или во влажном органическом субстрате, который дает им пищу. Чем ниже уровень эволюционного развития наземных растений, тем в большей сте пени они еще зависят от воды. У более прими тивных растений (например, плаунов, хвощей, особенно мхов) некоторые фазы развития орга низма происходят только в воде. Но приспосаб ливаясь к условиям суши, растения «избрали» альтернативную эволюционную стратегию. Суще ствование наземного растения зависит от почвен ной воды, и чтобы эта вода стала доступной, рас тение должно было развить корневую систему, поглощающую воду из почвы. У водорослей кор ней нет, так как они им не нужны, некоторые из них имеют «пятки», прикрепляющие их к кам ням, но ничего не всасывающие. Первые назем ные растения не имели корней, у них были волоскоподобные органы — ризоиды, которые про никали в почву и всасывали воду. Однако окон чательно обосновавшись на суше, растения раз вили корни.
Корни наземных растений всасывают из поч вы не только воду, но и минеральные соли, необ ходимые для питания растений. У водорослей и в данном случае не возникает проблемы, так как источником минеральных веществ для них слу жит вода, которая их омывает и представляет сво его рода питательный бульон, содержащий все необходимое для жизни. Переселившись на сушу, растения должны были найти какой-то другой способ снабжения минеральной пищей, поэтому их корни стали поглощать не только воду, но и минеральные вещества. Таким образом, корни на земных растений закрепляют их в грунте и по глощают воду, необходимую для поддержания формы тела и фотосинтеза, а стебли служат опо рой основным фотосинтезирующим органам — листьям. Вода непрерывно поступает в корневые волоски, движется вверх по корням и стеблям, а затем выводится наружу листьями. Все наземные части растения, в конечном счете участвующие в фотосинтезе, покрыты кожицей и восковидной ку тикулой, снижающей потери воды, но одновре менно затрудняющей необходимый газообмен между растением и окружающим воздухом. Эта проблема решается с помощью специальных от верстий — устьиц, которые открываются и закры ваются, регулируемые внешними и внутренними факторами, помогая таким образом поддерживать нужное соотношение между расходом воды и по требностями в углекислом газе и кислороде.
Возникновение эпидермиса в процессе эво люции наземных растений в конечном счете ре-
349
шило проблему их выживания: водные растения, оказавшись вне воды, быстро гибнут от высыха ния. Эпидермис возник в результате специализа ции наружных клеток тела растения.
Наличие устьиц является одним из наиболее характерных признаков высших растений, начи ная с древнейших известных их представителей — риниофитов.
Наиболее примитивным среди высших расте ний с эволюционной точки зрения является аномоцитный тип устьичного аппарата, обнаружен ный у риниофитов. Эволюция устьичного комп лекса связана со способом происхождения из ма теринской клетки (онтогенетические типы устьиц) либо с постепенной дифференциацией клеток, окружающих устьичную щель, увеличением их количества (морфологические типы устьиц).
Развитие вокруг устьичных щелей специаль ных клеток (замыкающих, побочных) разрешило проблему излишнего испарения воды через усть ица, так как устьица благодаря этому закрывают ся на ночь, когда из-за отсутствия света процесс фотосинтеза прекращается.
Основной орган фотосинтеза, лист, стал пло ским и тонким, так что клетки, в которых обра зовывался крахмал, получали весь свет, необхо димый для этого процесса. Развилась листовая мозаика, в листьях возникла эффективная систе ма внутренней вентиляции.
Уоднолетних растений фотосинтезирующим органом является также стебель. У многолетних он часто утолщается, одревесневает и покрывает ся пробкой, которая, как и кутикула, снижает потери воды.
По мере того как растения «поднимались» над землей, вставала проблема передачи воды и ми неральных веществ от корней к самым верхним ветвям, а продуктов фотосинтеза — от листьев к самым нижним частям корней. Так, хвойное де рево секвойя вечнозеленая (Sequoia sempervirens) достигает в высоту 110—112 м, и так высоко должна подняться вода и так низко должны опус титься пластические вещества. Транспорт раз нообразных веществ между фотосинтезирующими и нефотосинтезирующими частями растений происходит по стеблю и особой проводящей сис теме тканей. В зачаточной форме проводящую систему можно наблюдать у некоторых крупных морских водорослей, где специальные клетки об разуют как бы трубку, по которой передвигается вода с растворенными в ней питательными веще ствами.
Увысших растений проводящая система со стоит из двух основных компонентов: ксилемы (древесины), по которой вода проходит вверх по растению, и флоэмы (луба), разносящей питатель ные вещества, образованные в листьях и других фотосинтезирующих органах, по всему его телу. В процессе эволюции высших растений элементы ксилемы и флоэмы претерпели значительные морфологические и функциональные изменения,
очем говорилось в соответствующих разделах книги. Наиболее эффективным водопроводящим элементом является сосуд, что и закреплено в на звании основной группы растений — сосудистые.
Наземные растения столкнулись со многими механическими проблемами, чуждыми водорос лям. Главная из них — необходимость стоять прямо, тогда как тело водорослей поддерживает вода. Теоретически предела, которого могут дос тигать размеры водорослей, не существует, и длина, например, крупной бурой водоросли макроцистис (Macrocystis pyrifera) может достигать более 180 м. У наземных растений дело обстоит иначе. Первые наземные растения от земли не отрывались, стелясь по ее поверхности. «Подни маясь» вверх, растение должно было иметь доста точно прочный стебель, чтобы не только поддер живать ветви, но и противостоять ветру и другим механическим воздействиям. На первых порах эту проблему разрешило возникновение и развитие ксилемы, ее расположение в стебле и корне, поз воляющее стеблю сопротивляться изгибу, а кор ню — противостоять растяжению, сохраняя в то же время максимальную гибкость. У первых на земных растений был простой древесинный ци линдр, проходящий в центре стебля (протостела), он обеспечивал проведение воды и придавал прочность стеблю. По мере увеличения размеров
иусложнения строения растений шла эволюция протостелы, появились другие типы стелы, наи лучшим способом удовлетворявшие нужды расте ния. Произошла дифференциация и самих кле ток, составляющих древесину: одни из них (трахеиды, сосуды) стали выполнять единствен ную функцию — проведение воды, другие -
склеренхимные волокна, с сильно утолщенными и одревесневшими оболочками, приняли на себя обязанность укрепления стебля. В процессе эво люции у наземных растений постепенно возникла группа механических, опорных тканей, придаю щих прочность органам на изгиб или излом
(древесинные и лубяные волокна, располагающиеся вдоль осевых органов) либо на растяжение и сжа тие (колленхима, склереиды, располагающиеся или под эпидермисом, или внутри сочных паренхиматизированных органов).
Вотличие от животных растения растут всю жизнь. Их рост обеспечивается меристемами — ограниченными участками тканей, постоянно со храняющих эмбриональное состояние. Верхушеч ные (апикальные) меристемы, расположенные на концах всех корней и побегов, обеспечивают пер вичный рост — удлинение растений. В результате корни все время отыскивают новые источники воды и минеральных веществ, а фотосинтезирующие органы всегда тянутся к свету. Вторич ный рост обусловлен боковыми (латеральными)
меристемами—камбием и пробковым камбием (феллогеном) и приводит к утолщению стеблей, ветвей
икорней.
Меристемы обусловливают также ветвление растений, обеспечивающее постоянное увеличе ние фотосинтезирующей поверхности.
Предполагают, что высшие растения произо шли от водорослеподобных предков, обладающих
дихотомическим (изотомия) ветвлением талломов. Поэтому такой тип ветвления осевых органов на земных растений считается первичным. Он со хранился у некоторых более примитивных выс-
350
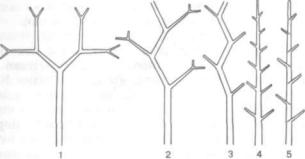
Рис. 315, Схема эволюции ветвления высших растений:
/ — изогамия (равная дихотомия); 2 — анизогамия (неравная дихотомия); 3 — дихоподай; 4 — моноподий; 5 — симподий
ший растений — плаунов, некоторых папоротни ков (рис. 315, 1). В процессе эволюции у высших растений появилось боковое ветвление, что можно рассматривать как проявление одной из общих закономерностей эволюции живых существ — дифференциации. При боковом ветвлении глав ная и боковые оси становятся неравноценными по структуре и функциям.
Врезультате неравного роста двух боковых ветвей возникла анизотомия (греч. anisos — не равный и tome — сечение). Она считается пере ходным типом между дихотомией и боковым ветвлением. Такое перерастание (перевершинивание) ветвей хорошо выражено у примитивного вымершего девонского рода хорнеофит (Ногпеоphyton; рис. 315, 2).
Дальнейшим шагом в эволюции ветвления яв ляется дихоподиальное ветвление, при котором две боковые оси развиваются резко неравномерно, одна из них перевершинивает другую (А. Л. Тахтаджян, 1978). Дихоподиальное ветвление было характерно для примитивного девонского рода астероксилон (Asteroxylon); оно имеет место у многих плаунов и селагинелл, у некоторых древ нейших хвощевидных и у многих папоротнико видных (рис. 315, 3). Ряд авторов относят дихо подиальное ветвление к симподиальному.
Врезультате одного или нескольких ветвле ний образуется система осей (корней, побегов), находящихся между собой в той или иной степе ни подчинения.
Большинство ботаников основные типы сис тем осей при боковом ветвлении относит к моноподиальным и симподиальным.
Всоставе моноподиальной системы каждая ось представляет собой моноподий (греч. monos — один, единый и pus, род. п. podos — нога), т. е. результат работы одной верхушечной меристемы (рис. 315, 4). Основное отличие моноподиального ветвления от дихотомического и дихоподиального заключается в том, что боковые ветки закладыва ются под верхушкой главной оси.
Из моноподиального ветвления в процессе эво люции возникает симподиальное (греч. sym — вме сте). В составе симподиальной системы форми руются симподии, или составные оси, как резуль тат перевершинивания (рис. 315, 5). У высших растений моноподиальными и симподиальными могут быть и корневые системы, и системы побе гов.
У высших растений особенно четко проявился особый аспект дифференциации тела, который называют полярностью.
Полярность (лат. poms, греч. polos — полюс) — свойственная организмам специфическая ориен тация процессов и структур в пространстве, при водящая к возникновению морфофизиологических различий на противоположных сторонах (или концах) клеток, тканей, органов и организма в целом. У растений полярность проявляется осо бенно четко. Так, еще у многих одноклеточных водорослей на нижней (затененной) стороне об разуются корнеподобные выросты — ризоиды, а на верхней (освещенной) — органы фотосинтеза, которые, однако, могут быть переориентированы путем затенения верхней и освещения нижней стороны клетки (например, каулерпа). У много клеточных низших растений полярность выраже на сильнее и является более фиксированной. Так, у зеленых водорослей она проявляется в том, что каждая клетка способна при определенных усло виях образовывать в своей морфологически ниж ней части ризоиды, а в верхней — фотосинтезирующий орган.
Наибольшей сложности полярность достигает у высших растений, приспособленных к жизни в воздушно-почвенной среде. В таком случае она проявляется в расчленении тела на корни и по беги, в различии между основанием и верхуш кой отдельного органа, в строении и функции разных тканей и клеток, в способности к регене рации.
У спор мхов, хвощей, папоротников поляр ность возникает после соответствующего внешне го воздействия, когда клетки начинают делиться, и дают начало новому организму. Первая перего родка в прорастающей споре ориентируется пер пендикулярно падающему лучу света, разделяя спору на затененную «корневую» и освещенную «заростковую» клетки. У высших семенных рас тений полярность наблюдается уже в зиготе, где формируются два зачаточных органа — побег с верхушечной почкой и корень. У формирующего ся растительного организма полярность проявля ется в направлении деления клеток, их роста и дифференциации; ведущая роль в этом процессе принадлежит фитогормонам. Многие вещества синтезируются в определенных тканях и переда ются по проводящим путям полярно. Например, синтезируемые в верхушке побега ауксины пере текают к его основанию, возбуждая работу кам бия, но подавляя вегетацию боковых почек.
Полярность сформировавшихся органов выс ших растений, как правило, сохраняется даже при резком нарушении их нормального положе ния. Это особенно наглядно проявляется при ве гетативном размножении. Какой бы стороной мы не поместили черенок смородины, тополя, ивы в почву, корни всегда развиваются на морфологи чески нижней, а побеги — на морфологически верхней стороне. Однако наблюдались случаи, когда резкие экстремальные факторы (изверже ния вулканов, морские штормы, землетрясения, высокая радиация) как бы «перетрясают» орга низм, вызывая сдвиги в проявлении полярности.
351