

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfРис. 228. Участок поперечного среза корневища пырея
(Agropyron re pens): |
|
) — эпидермис; 2 — паренхима коры; 3 — эндодерма; 4 - |
склеренхима пери- |
цикла; 5 — проводящие пучки |
|
находятся лишь чешуевидные мелкие листочки, соответственно формируется слабая проводящая система, в ксилемной части состоящая из редких узкополостных сосудов примитивного строения. После того как начнет развиваться надземный побег с нормальными листьями, повышается ак тивность камбия, увеличиваются число и просвет сосудов.
Подземные столоны и клубни. Клубень (tuber - видоизмененный побег, стебель которого, вклю чающий одно или несколько междоузлий, сильно разрастается и накапливает запасные вещества (крахмал, реже масла). Орган возобновления дву- и многолетних клубненосных растений, перенося щий неблагоприятный период (зимний, засушли вый), нередко служит для вегетативного размно жения.
Клубни бывают подземные и надземные. Под земные клубни часто возникают на столонах. Рассмотрим процесс формирования клубня у кар тофеля (рис. 229, А, Г)- Побеговая природа клуб ней обнаруживается по характеру расположения на них почек (глазков). Как и на обычных побе гах, расположение почек на клубнях спиральное, имеется верхушечная почка. Из глазков посажен ного в землю клубня образуются надземные об лиственные и цветущие побеги. В пазухах чешуе видных листьев, расположенных на подземных частях сформировавшихся побегов, летом вырас тают хрупкие белые столоны с недоразвитыми листочками. Столоны резко отличаются от прида-
Рис. 229. Клубни:
А — картофеля (Solanum tuberosum): I — прорастающий клубень (и — надземный побег, образующийся из верхушечной почки; б — глазки); 2 — клубнеобразование у картофеля; Б — поперечный срез клубня картофеля (схемат.): 1 — пробка; 2 — кора; 3 — проводящие пучки; 4 — паренхима; 5 — сердцевина; В — цикламен пурпуровый (Cyclamen purpurascens): a — клубень; Г — колоказия древняя (Colocasia antiquorum), или таро: клубни широко культивируемой формы «дашин»; Д — целогина гребенчатая с псевдобульбами; Е — кольраби (Brassica oleraceae L. var. gongyloides); Ж— диоскорея посевная, или ямс посевной (Dioscorea saliva): a — воздушные клубни; 3 — диоскорея слоновая (D. elephantipes); И— калатея Баше (Calathea bachemiana) с водозапасающими клубнями
252
точных корней, растущих рядом с ними на мате ринских стеблях, пазушным положением, толщи ной, наличием листьев-чешуек.
Верхушечные почки столонов начинают утол щаться за счет разрастания и паренхиматизации оси, превращающейся постепенно в клубень. Ли стья-чешуйки видоизменяются на поверхности клубня в бровки (листовой рубец), в пазухах их закладываются одна-три почки, так формируются глазки. Столоны быстро разрушаются, обособлен ные клубни служат органами вегетативного раз множения, что происходит в естественных усло виях у дикого картофеля на его родине в Андах. У культурного картофеля разрушение столонов и расселение клубней невозможно без помощи че ловека, но в основе приемов возделывания лежат естественные процессы клоноразмножения с по мощью подземных столонов и клубней.
Внутреннее строение клубня картофеля пока зывает, что это сильно паренхиматизированный подземный побег (рис. 229, а, 2). Общий план строения обнаруживает его стеблевое происхож дение. Снаружи клубень покрыт пробкой. Как в коровой, так и в сердцевинной паренхиме накап ливается крахмал. По камбиальному кольцу на большом расстоянии друг от друга расположены слаборазвитые проводящие пучки. Основную массу клубня составляет паренхима внутренней флоэмы и сердцевины.
Так же, как у картофеля, происходит клубнеобразование, возобновление и вегетативное раз множение у широко распространенного в сосно вых лесах седмичника европейского, стрелолиста, топинамбура.
Подземные клубни могут формироваться пу тем утолщения подсемядольного колена, как у цикламена (рис. 229, В); на корневищах путем его утолщения, как у колоказии сем. аронниковых (рис. 229, Г).
Надземные клубни бывают различного проис хождения. У некоторых лиан, например у церопегии Вуда (Ceropegia woodu) из сем. ластовненых (Asclepiadaceae), стеблевые клубни представляют утолщенные узлы вместе с основаниями двух су противных листьев. Эти клубни связаны между собой участками стебля и формируют придаточ ные корни, с помощью которых прикрепляются к опоре.
Умногих орхидных имеются надземные клуб ни, нередко называемые псевдобульбами. Это утолщенные участки стебля, верхушечная почка которого редуцирована, и несут только один или два нормально развитых листа. Новый побег об разуется из пазухи чешуи у основания клубня и заканчивается клубнем. Таким образом, получает ся симподиальный побег из ряда зеленых клуб ней, несущих на верхушке зеленые листья и свя занных тонким стеблем с чешуевидными листья ми (целогина гребенчатая, рис. 229, Д).
Ушироко известного растения кольраби глав ный стебель над поверхностью почвы клубневид но утолщен и несет на своей верхушке листья (рис. 229, Е).
Унекоторых представителей сем. диоскорейных (Dioscoreaceae) также образуются стеблевые
клубни. У тамуса обыкновенного они развивают ся из первого междоузлия над семядолями и мо гут достигать больших размеров. Из клубня еже годно возникают побеги. Клубни характерны для большинства видов рода диоскорея (Dioscorea), у которых наряду с однолетними и многолетними подземными клубнями, есть полунадземные и надземные (рис. 229, Ж). В сухих субтропиках Юго-Западной Африки на высоте 170—1300 м встречается другой представитель этого рода с многолетним полунадземным клубнем, возни кающим из зоны гипокотиля. Это знаменитая диоскорея слоновая, или слоновая нога, одно из чудес здешней своеобразной природы (рис. 229, 3). Огромный клубень мозаично одет снаружи толстыми и прочными многоугольными плитками из пробки. Из клубня каждую весну возникают побеги, отмирающие в сухой период.
У некоторых растений образование клубней связано с запасанием воды. Так, у представителей сем. марантовых (Marantaceae), распространенных
втропической зоне всех континентов, кроме Ав стралии, корневища образуют направленные в глубь почвы утолщенные столоны, в которых на капливается вода. Такие клубневидные столоны образует маранта тростникововидная, виды калатеи (рис. 229, И). Мелкие клубеньки могут воз никать в области соцветия (горец живородящий)
ввиде выводковых почек.
Вприроде существует много переходных форм между толстыми корневищами и клубнями, на пример у норичника, чистеца болотного, мяты полевой.
Луковица (bulbus). Это подземный, реже над земный побег с коротким уплощенным стеблем — донцем — и мясистыми сближенными чешуевид ными листьями, запасающими воду и раствори мые питательные вещества, преимущественно са хара. На донце находится верхушечная почка, а в пазухах чешуи — боковые, из которых вырастают надземные побеги, от донца в почву отходят при даточные корни (рис. 230, А).
Луковичные растения принадлежат почти ис ключительно к однодольным, в основном к сем. лилейных (Liliaceae) и амариллисовых (Amaryllidaceae). Как исключение, луковицы встречаются
удвудольных, например у некоторых видов рода кислица (рис. 230, Б).
Число чешуи в луковице может сильно варьи ровать. Так, луковица может иметь только одну чешую (кандык сибирский), две-четыре (виды рода рябчик), много (виды родов нарцисс, про лески).
По строению и расположению чешуи бывают трех основных типов: черепитчатые — чешуи низкие и соприкасаются только краями или лишь слегка прикрывают рядом расположенные (рис. 230, В, а, б); концентрические — каждая чешуя, срастаясь краями, полностью охватывает после дующую, в результате чего чешуи вложены одна в другую (рис. 230, Г, а, б); полуконцентрические —
чешуи, охватывая наполовину или более после дующую, не срастаются своими краями. Между основными типами чешуи имеются переходы.
Природа луковичной чешуи бывает различна.
253
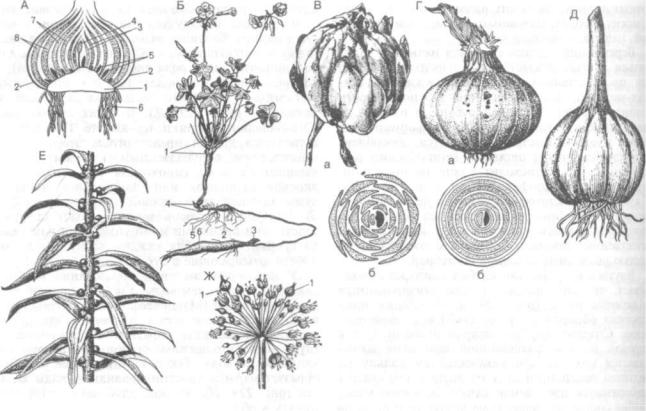
Рис. 230. Луковицы:
А — строение луковицы (схемат.): 1 — донце; 2 — отмерите чешуи; 3 — листья будущего вегетационного перехода; 4 — зачаток цветка; 5 — почки; 6 — придаточные корни; 7 — чешуи — листовые влагалища; 8 — чешуи — низовые листья; Б — кислица козья (Oxalis pes-caprae): I — пазушные луковички в основании черешков листьев; 2 — материнская луковица подземного побега; 3 —- почка возобновления; 4 — дочерние луковички; 5 — придаточные корни (всасывающие); 6 — контрактиальный (втягивающий) корень; В — черепитчатая (чешуйчатая) луковица лилии Кессельринга (Lilium kesselringianum): a — общий вид; б — схема поперечного среза; Г — лук репчатый (Allium сера) с концентрическими чешуями: а — общий вид; б — схе ма поперечного среза; Д — сложная луковица чеснока (Allium sativum); £ — лилия луковичконосная (Lilium bulbiferum) с луковичками в пазухах листь ев; Ж— луковички (1) в соцветиях лука (Allium sp.)
У одних растений чешуи представляют собой мя систые низовые листья, которым свойственна только функция накопления питательных веществ (лилия-саранка, виды тюльпанов), у других — че шуи являются разросшимися мясистыми основа ниями ассимилирующих листьев, которые утол щаются и остаются в составе луковицы после от мирания зеленых пластинок (лук репчатый, лук медвежий). Луковица может также состоять из чешуи обоих типов (нарцисс).
В некоторых луковицах различают еще влага лищную чешую. Она закладывается в онтогенезе луковицы первой, имеет вытянутую замкнутую верхушку и охватывает молодую почку. При про растании луковицы ее верхушка прорывается, чешуя постепенно отмирает.
Донце луковицы может быть довольно долго вечным и состоять из годичных приростов двух, трех и более лет (у лилий) или же луковица еже годно сменяется, представляя собой одногодич ный побег (некоторые виды луков).
По способу нарастания луковицы могут быть моноподиальными — возобновление идет из вер хушечной почки, а цветоносные побеги образу ются из пазушных почек (нарцисс, подснежник, рис. 231, А) и симподиальными — цветоносный побег развивается из верхушечной почки, а во зобновление происходит из пазушных (тюльпан, рябчик, лук, гиацинт; рис. 231, Б). Наружные чешуи, израсходовав запасы питательных ве ществ, подсыхают и играют защитную роль.
Если в рост трогаются две или несколько по чек, луковица ветвится, образуя луковицы-детки. У сложных луковиц (чеснок, см. рис. 230, Д) в пазухе каждой чешуи формируется несколько лу ковиц-деток — зубков.
Систематическими признаками луковиц яв ляются их форма (круглая, яйцевидная, конусо видная, продолговатая, веретеновидная, неравнобокая, сплюснутая и т. д.), размеры, окраска на ружных чешуи (от белой до темно-коричневой).
Подземные луковицы как органы запаса и во зобновления свойственны главным образом эфе мероидам сухих жарких областей (виды тюльпа на, лука), высокогорья (луки, рябчики) и широ колиственных лесов (подснежник, пролеска и др.).
Помимо подземных, существует ряд луковиц, которые образуются в пазухах листьев надземного побега. Они обычно называются луковичками (bulbilli), возникают из пазушных почек и служат для вегетативного размножения, например у зубянки луковичной, лилии луковичконосной (см. рис. 230, Е). Луковички легко отделяются от стебля и, падая на землю, быстро укореняются.
У многих растений луковички возникают в соцветиях: некоторые виды лука (см. рис. 230, Ж), агавы, мятлик альпийский и др.
Клубнелуковица (bulbotuber). Внешне клубне луковица напоминает луковицу, но накапливает питательные вещества в разросшемся мясистом стебле подобно клубню. Чешуевидные листья су хие, пленчатые, часто это просто остатки влага-
254

Рис. 231. Нарастание оси луковицы:
А — моноподиально нарастающая многолетняя луковица нарцисса (Narcissus); Б — симподиально нарастающая многолетняя луковица гиацинта (Hyacinthus); 1 — донце (укороченный, многолетний стебель); 2 — отмершие че шуи; 3 — листья будущего вегетационного периода; 4 — зачаток цветка; 4а — отмерший цветок; 5 — почка возоб новления (боковые не показаны); 6 — придаточные корни; 7 — чешуи — низовые листья; 8 — чешуи — листовые влагалища; 9 — остаток цветоноса текущего года; 9а — остатки цветоносов предыдущих лет, 1992—1995 — годич ные циклы соответствующих лет
лищ отмерших зеленых листьев, выполняют толь |
1962), но чаще каудексом (лат. caudex — ствол, |
ко защитную функцию. Таким образом, клубне |
стержень). Каудекс вместе с корнем выполняет |
луковица — это скорее облиственный клубень, а |
функцию запаса питательных веществ и образует |
не луковица. Характерна для гладиолусов, без |
большое количество почек возобновления. В со- |
временников, шафранов (рис. 232). |
|
Каудексы. У многолетних трав и полукустар |
|
ничков с хорошо развитым, сохраняющимся всю |
|
жизнь стержневым корнем формируется своеоб |
|
разный многолетний орган стеблекорневого про |
|
исхождения, который некоторые морфологи на |
|
зывают стеблекорнем (Ал. А. Федоров и др., |
|
Рис. 232. Клубнелуковицы: |
Рис. 233. Каудексы: |
|
А — шафран посевной (Crocus sativus): a — общий вид, б — поперечный срез |
1 — ветвистый мелколепестника кабульского (Erigon cabulicus); 2 — ежовник |
|
щетинко-волосый (Anabasis hispidula): a — почки возобновления; 3 — погру |
||
(схемах); 1 — укороченный стебель; 2 — отмершие чешуи; 3 — листья буду |
||
женный горца дубильного (Polygonum coriarium); 4 — начало партихуляции |
||
щего вегетационного периода; 4 — зачаток цветка; 5 — почки; 6 — придаточ |
||
каудекса у василька шероховатого (Centaurea scabiosa): a — основания цвету |
||
ные корни; 1994—1995 — годичные циклы соответствующих лег, Б — гладио |
||
щих побегов |
||
лус (Cladiolus) |
||
|
255
став каудекса могут входить основания отмерших цветоносных побегов, втягивающиеся в почву укороченные оси розеточных побегов, одревесне вающий гипокотиль, разросшаяся корневая шей ка. В стеблевой части каудекса вначале хорошо заметны листовые рубцы, определенная законо мерность в расположении почек, которая позже может быть замаскирована, так что порой трудно отличить, где кончается побеговая часть каудекса и начинается стержневой корень. Стержневые корни, на которых формируется каудекс, облада ют конграктильностью, способностью втягивать растение в почву.
Каудекс особенно характерен для полупус тынных, пустынных и высокогорных полукустар ничков, представителей сем. розоцветных, астро вых, маревых, свинчатковых. В нашей зоне он наблюдается у василька шероховатого, свербиги, клевера горного, люпина многолистного, люцер ны, полыни, одуванчика и др. Нередко каудекс достигает значительных массы и размеров; на пример, у некоторых видов рода прангос (Ргапgos) он весит до 15 кг.
Каудекс бывает простой и ветвистый, надзем ный и подземный, или погруженный (рис. 233). Напоминая внешне короткие толстые корневища, он отличается от них способом отмирания — от центра к периферии (как в стебле). В централь ной части каудекса образуется полость, а за тем каудекс вместе с корнем часто делится на от дельные участки — партикулы (лат. partis - часть).
Процесс разделения каудексовых растений на части называется партикуляцией. Отделившиеся части растения представляют собой старческие особи, не способные цвести и плодоносить (сенильные растения; лат. senilis — старче ский).
Метаморфоз органов растений давно привле кал пристальное внимание естествоиспытателей. Детальные исследования метаморфоза провел И. В. Гёте, известный натурфилософ и еще более известный немецкий поэт. Позднее метаморфоз у растений и животных изучал Ч. Дарвин (о теории метаморфоза см. заключительную главу).

Глава
Размножение
растений
ОБЩЕЕ ПОНЯТИЕ О РАЗМНОЖЕНИИ И ЕГО ФОРМАХ
Размножение — это присущее всем живым ор ганизмам свойство воспроизведения себе подоб ных, обеспечивающее непрерывность и преемст венность жизни. Результатом размножения является длительное существование видов в пространстве и во времени. Потеря способности к воспроизве дению ведет к вымиранию вида.
Иногда размножение трактуют как увеличение числа особей данного вида. Однако вид может существовать неопределенно долго, не увеличи вая числа особей, а заменяя дочерними организ мами родительские. Такое явление часто называ ют воспроизведением.
В узком смысле размножение можно понимать как появление новых особей со всеми особенно стями родительского организма, что наблюдается при размножении частями тела. Результатом же полового процесса является появление новых особей, отличающихся от исходного организма, что дает основание говорить о воспроизведении.
Вряд ли есть смысл искать противоречия меж ду этими понятиями. Цель размножения — полу чить новые организмы путем их воспроизводства любым способом для обеспечения существования вида. При этом число новых особей может быть больше, чем родительских, равно или меньше. Нельзя забывать, что численность потомства в значительной степени зависит от внешних усло вий.
Различают два пути размножения — половой и бесполый. Сущность полового размножения за ключается в том, что дочерние особи развиваются из зиготы, образующейся в результате слияния женской и мужской половых клеток — гамет.
<зГ+ |
2п |
митоз -* дочерний организм |
Гаметы генетически различны, что ведет к увеличению изменчивости потомства и создает благоприятные условия для естественного отбора. Этот путь размножения имеет многообразные формы, обусловленные размножением, подвиж ностью гамет, различными отклонениями от нор мального пути.
Бесполое размножение характеризуется отсут ствием полового процесса и осуществляется без участия половых гамет. Представлено двумя фор мами: а) размножение делением родительской особи на более или менее равные части (у одно клеточных водорослей), или делением вегетатив ного тела на участки разной величины (черен кование листа, стебля, корня), или обособлением его частей. Многие растения формируют вегета тивные зачатки: почки, клубеньки, луковички. Такая форма бесполого размножения называется
вегетативным размножением; б) размножение специализированными клетками — спорами (греч. spora — посев, сеяние) является древнейшей фор мой размножения у одноклеточных организмов, но имеет место и у многоклеточных — грибов, растений. У высших растений формирование спор связано с редукцией числа хромосом, след ствием чего является морфологическое различие между дочерними и родительскими растениями.
Обе формы бесполого размножения осуществ ляются многочисленными способами. В процессе онтогенеза размножение может быть однократ ным (организмы с однократным размножением называются моноциклическими и приносят, как правило, многочисленное потомство) или много кратным (такие организмы называются полицик лическими и обычно менее плодовиты). У расте ний в жизненном цикле наблюдается смена по лового (гаметофит) и бесполого (спорофит)
17 Зах. 1851 |
257 |
поколений. Один и тот же организм может раз множаться как половым, так и вегетативным пу тем, что и обеспечивает растениям способность к быстрому и широкому расселению.
ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ РАСТЕНИЙ
Вегетативное размножение — образование но вой особи из родительской. Жизнеспособная часть вегетативного тела растения, естественно или искусственно отделенная от родительской особи, образует недостающие органы и развива ется как самостоятельный организм. Таким обра зом, в основе вегетативного размножения расте ний лежит широко распространенная у них спо собность к регенерации (лат. regeneratio. — воз рождение, обновление), т. е. возобновление утра ченных органов, частей или развитие из отдель ных участков тела вновь всего растения. При этом все необходимые органы у будущей особи могут формироваться как после отделения от ро дительского растения, так и до обособления (на пример, выводковые побеги бриофиллюма, розеточные побеги с придаточными корнями на кон цах усов земляники).
Способность размножаться вегетативно харак терна для растений разных уровней организации (водоросли, грибы, лишайники, высшие споро вые и цветковые растения), при этом вегетатив ное размножение может сопутствовать семенному (половому) или, наоборот, может преобладать (у осины, ив, малины), а у некоторых оно может быть исключительным, как у элодеи канадской. В Европу элодея завезена только в форме женских экземпляров, и поэтому она размножается только вегетативно. У многих хвойных эта способность выражена слабее, а у некоторых в естественных условиях вообще не наблюдается (сосна обыкно венная).
Способность к вегетативному размножению не остается одинаковой в процессе онтогенеза. Известно, что с возрастом резко снижается спо собность к образованию пневой поросли у дре весных растений, что учитывается работниками лесного хозяйства при определении возраста руб ки насаждений. Это очень важно для ведения хо зяйства в дубравах, грабовых, березовых и ольхо вых лесах.
Совокупность особей, возникающих из одной родительской особи в результате вегетативного
размножения, образует клон (греч. clon — от прыск, ветвь). Наглядно проследить формирова ние клонов можно у осины, земляники, терна, малины.
Вегетативное размножение имеет как положи тельные, так и отрицательные стороны. Оно по зволяет в полной мере сохранить у потомков ка чества родителей, что очень важно для сохране ния чистых линий. Эта особенность широко используется в практике садоводства, селекции. Кроме того, оно практически не зависит от кли матических условий, которые в некоторые годы полностью исключают образование семян. Одна
ко многократное воспроизведение особей вегета тивным путем ведет к накоплению возрастных изменений, увеличению общего возраста орга низма. При этом происходит старение фермента тивных систем, белков, снижаются продолжи тельность жизни, устойчивость к вредителям и болезням. Примером тому могут служить пира мидальные тополя, широко применяемые в озе ленении на территории Беларуси. К 40—50 годам они начинают усыхать и отмирают, что является результатом многовекового вегетативного раз множения мужских особей.
Принято различать естественное и искусст венное вегетативное размножение, хотя провести четкое разграничение между ними очень трудно.
ЕСТЕСТВЕННОЕ ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ
Под естественным вегетативным размножени ем понимают размножение вегетативными орга нами или их частями, происходящее в природе без вмешательства человека. Оно осуществляется различными способами (рис. 234).
Размножение делением клетки у одноклеточных водорослей или случайным отделением от таллома отдельных участков у многоклеточных водорослей, грибов, лишайников. В результате чрезвычайной способности к регенерации эти участки развива ются в самостоятельные растения. Очень часто при разрушении старых частей таллома более мо лодые его ветви обособляются и становятся само стоятельными организмами. Так могут размно жаться и цветковые растения. Ряски, например, из нескольких перезимовавших особей дают по томство, занимающее площади в сотни квадрат ных метров. Таким же образом размножаются и элодея канадская (канадская чума), многие ивы. Обычно после паводка по берегам рек можно увидеть много укоренившихся побегов ив.
Выводковые почки — это специализированные придаточные почки, которые дают начало новым особям. Формирование выводковых почек харак терно как для слоевцовых растений (водоросли, печеночники), так и для высших споровых и цветковых. Образуются они на жилках листьев (папоротник асплениум) или на хлыстовидно от тянутых верхушках листьев (так называемый странствующий папоротник из сем. Асплениевых — Camptosorus rhizophullus); в пазухах листь ев (чистяк, некоторые виды лилий); по краю лис та (у бриофиллюма — Bryophyllum). По краю зуб чатого листа бриофиллюма в каждой выемке формируется придаточная почка, здесь же про растающая в побег (2—3 листочка и 1—2 прида точных корешка). Осыпаясь, они легко укореня ются, давая массовый «самосев». У зубянки (Dentaria) выводковые почки видоизменены в луковички, а у горца живородящего — в стебле вые клубеньки.
Выводковыми почками часто называют не большие вегетативные побеги, образующиеся в пазухах листьев, в соцветиях, укореняющиеся при опадении. Это явление свойственно для растений полярных районов, высокогорий и степей — мят-
258

Рис. 234. Способы вегетативного размножения:
1 — выводковые почки на листе Bryophylluma; 2 — часть листа с выводковыми почками папоротника Asplenium; 3 — зимующие почки у стрелолиста; 4 — соцветие с выводковыми почками у мятлика луковичного (Роа bulbosa): a — об щий вид соцветия, б — выводковая почка; 5 — ползучие побеги (усы) у земляники; 6 — корневые отпрыски горной яб лони
лик степной (Роа bulbosa), некоторые камнеломки (Saxiftaga), очитки (Sedum), арктические овсяни цы (Festuca). Такие растения называют живоро дящими, хотя и не совсем точно. Истинное жи ворождение характерно для мангровых растений и выражается в прорастании семян на материн ском растении. Молодое растеньице падает на грунт в момент отлива и ко времени следующего прилива достаточно прочно закрепляется в суб страте.
Зимующие почки образуются на вершинах стеблей или на особых боковых побегах у многих водных, главным образом плавающих, растений. Зимующие почки — гибернакулы (лат. ЫЬёгпшп — зима) формируются у пузырчатки, телореза, ля гушатника, урути, некоторых рдестов. Осенью в них накапливается крахмал и они вместе с мате ринским растением или отделившись от него опускаются на дно. Весной, отделившись от пере гнивших растений, зимующие почки всплывают (так как имеют воздухоносные полости) и разви ваются в новые растения.
Размножение при помощи надземных ползучих побегов (плети, усы, столоны) происходит у зем ляники, костяники, некоторых лапчаток, живуч ки ползучей, будры и др. Годичный прирост каж дой плети может достигать 1,5 м (у земляники), а число новых особей в течение двух лет увеличи вается до 200 и более. Надземные ползучие побе ги экологически представляют собой переход от вертикальных стеблей к корневищам. Стелясь по поверхности субстрата, они в узлах образуют при даточные корни и здесь же, в пазухах листьев — почки, из которых развиваются вертикальные об лиственные побеги. После отмирания междоуз
лий ползучих побегов новые растения теряют связь с материнским.
Размножение корневищами наблюдается у большинства многолетних трав. На коротких корневищах почки сближены, поэтому надземные побеги, развивающиеся из них, скучены. На уд линенных корневищах почки достаточно удалены друг от друга, поэтому надземные побеги рассре доточены. При отмирании корневища новые осо би существуют самостоятельно. Ежегодный при рост корневищ составляет у ветрениц и душицы 5—10 см, у тысячелистника, полевого хвоща — 10—15, пырея ползучего — до 30, иван-чая — 85— 100, а у сахалинской гречихи — 150—300 см. Раз множение корневищами сохраняет почти неиз менным видовой состав лугов, скашиваемых во время цветения злаков, которые из-за этого не могут размножаться семенами.
Размножение луковицами характерно для мно гих травянистых, большей частью однодольных растений (луки, чеснок, тюльпаны, гиацинт, нар цисс, лилии и др.).
Луковицы образуются под землей (тюльпан, луки), в пазухах надземных стеблей (зубянка) или в соцветиях (чеснок). Дикий чеснок способен образовывать до 600 кг луковичек (для сравне ния — при посеве пшеницы на 1 га высевают до 100 кг зерновок).
Размножение клубнями происходит у неболь шого числа растений (картофель, земляная груша, кувшинка и др.). Клубень — это однолетний под земный побег растения с утолщенным стеблем, часто сферической формы. О стеблевой природе клубней свидетельствуют пазушные почки, из которых на следующий год вырастают новые по-
259
беги. Клубни могут быть и корневого происхож дения (георгин) — подземные и надземные.
Размножение корневыми отпрысками очень распространено в природе у дикорастущих расте ний и широко используется в практике лесовод ства и агролесомелиорации. При нанесении ран на корнях образуется большое количество прида точных почек, из которых развиваются новые особи. Такие растения очень ценны для облесе ния эродированных склонов, оврагов, восстанов ления леса после рубки. Хорошо развита способ ность образовывать корневую поросль у айвы, терна, маслины, розы, сирени, осины, боярыш ника, осота полевого и других. Поросль может быть не только корневой, но и пневой. Из спя щих почек в основании стебля развивается мощ ная поросль, укореняющиеся экземпляры кото рой способны заменить срубленный материнский ярус.
Способность формировать отводки в естест венных условиях обнаруживают пихты, липы, крыжовник. В местах соприкосновения с почвой нижние ветви образуют придаточные корни и начинают существовать как самостоятельные ор ганизмы.
ИСКУССТВЕННОЕ ВЕГЕТАТИВНОЕ РАЗМНОЖЕНИЕ
В узком смысле под искусственным вегета тивным размножением понимают такое, которое в природе не происходит, так как связано с хи рургическим отделением от растения частей для размножения. Искусственное вегетативное раз
множение используют, если растение дает мало семян или вообще их не образует, если при се менном размножении не сохраняются свойства сорта (у гибридов) или если необходимо быстро размножить сорт.
Использование человеком для размножения клубней, луковиц-деток, других способов естест венного размножения хотя и применяется очень широко, но не является искусственным. Наибо лее применимы следующие способы вегетативно го размножения (рис. 235): деление кустов, раз множение отпрысками и отводками.
Деление кустов применяют при размножении декоративных многолетников (примула, марга ритки, флоксы, дельфиниумы и др.), реже — у многолетних овощных (ревень, щавель, лук-ба- тун) и у некоторых кустарников (смородина, крыжовник). Куст (совокупность побегов) выка пывают из земли, расчленяют на особи с собст венными корнями и пересаживают на новое ме сто.
При использовании отпрысков материнское растение из земли не извлекается. Выкапыванием
ипересаживанием отпрысков размножают мали ну, ежевику, сливу, вишню, облепиху и т. д. Стеблевыми отпрысками размножают землянику
иклубнику, отделяя укоренившиеся растения. Отводки получают, пригибая нижние ветви
или стебли дугообразно к земле и засыпая их так, чтобы верхушка осталась на поверхности почвы. Для лучшего укоренения нижнюю часть ветви надрезают и при помощи приколов прижимают к субстрату. Надрез препятствует оттоку ассимилятов и способствует образованию каллюса и при даточных корней. Таким способом размножают
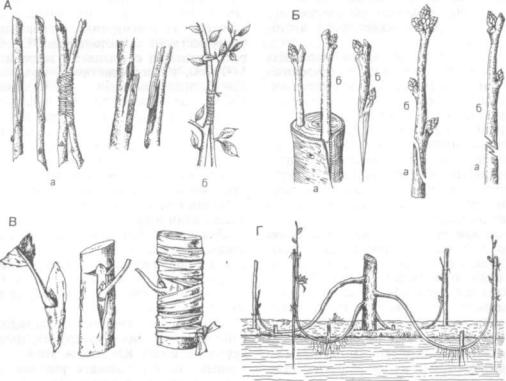
Рис. 235. Искусственное вегетативное размножение:
А — прививка сближением вприклад (а) и язычком (б); Б — прививка копулировкой (а — подвой; б - привой); В — прививка спящей почкой; Г— размножение отводками
260
крыжовник, шелковицу, лещину, виноград, аза лии и другие растения.
Черенкование как способ размножения в прак тике имеет исключительно большое значение. Черенок — это искусственно отделенный не большой побег или отрезок иного органа, взятый в целях его укоренения. Черенки бывают стебле вые, листовые, корневые. В зависимости от при роды исходного материала стеблевые черенки де лят на зимние (безлистные) и летние (с листьями, зеленые).
Зимние черенки готовят обычно из побегов данного года после их вступления в период зим него покоя. Длина черенков 20—30 см, зимой их хранят или в песке, или под снегом, весной вы саживают в субстрат. При размножении черенка ми необходимо учитывать явление полярности, т. е. специфическую ориентацию процессов и структур в пространстве, приводящую к форми рованию морфологических градиентов. Проявля ется полярность в процессе регенерации: у стеб левых и корневых черенков независимо от их по ложения в пространстве побеги развиваются с морфологически апикального (по отношению к вершине растения), а корни — с базального кон цов. Объясняется это явление тем, что индолилуксусная кислота (ИУК), перемещаясь полярно, скапливается в морфологически нижнем конце черенка и индуцирует включение генетической программы корнеобразования. Эту особенность растений необходимо учитывать при размещении черенков в грунте.
Летние черенки нарезают из облиствленных стеблей текущего года длиной 3—4 см с усло вием, что каждый черенок содержит один-два междоузлия. За два-три дня до использования побега на черенки листья укорачивают наполови ну, черенки помещают наклонно во влажный пе сок в парничок или теплицу, где создают высо кую относительную влажность, чтобы свести до минимума транспирацию. В местах поранения срезов из живых клеток образуется каллюс, в ко тором закладываются придаточные корни.
Черенкованием широко размножаются тополя, ивы, смородина и многие другие цветковые рас тения.
Листовой черенок — это удаленный с растения лист. У него лучше происходит формирование корней, чем у стеблей. Этот способ широко при меняют в цветоводстве при размножении бего ний, колеусов, гиацинтов, лилий, глоксиний и др. Предварительно надрезанный по крупным жилкам лист укладывают нижней стороной на влажный песок во влажной камере (под стеклян ный колпак). На нижней стороне формируются придаточные корни, на верхней — почки.
Корневые черенки представляют собой отрезки корней, которые заготавливают осенью и хранят во влажном песке. Длина черенков 10—20 см, в почву их укладывают наклонно и неглубоко заде лывают землей. Камбий и паренхима формируют каллюс, в котором образуются почки. Корневыми черенками размножают цикорий, вишню, сливу, иргу, орешник, малину, бересклет и многие дру гие растения.
Прививки (трансплантация) — это пересадка одного растения на другое с последующим их срастанием. Прививаемый компонент называют привоем, а растение с корневой системой — под воем. Прививки применяют как с целью получить новые сорта или изменить существующие («метод ментора», предложенный И. В. Мичуриным), так и с целью размножить имеющийся сорт с макси мальным сохранением его качеств. Существуют разные способы прививки.
Сближение, или аблактировка, применяется для рядом растущих растений. На обоих побегах срезают лентовидные участки коры (одинаковые по ширине и длине), затем побеги смыкают друг с другом и плотно обвязывают, оставляя на все лето и зиму. На второй год после срастания при вой отделяют от материнского растения, а у под воя удаляют верхушку.
Прививка черенком (копулировка) широко при меняется как в садоводстве, так и в лесоводстве при создании семенных плантаций. Черенок го товят из однолетних побегов, причем срезают бы стро, в один прием, косо. Срезанный черенок соединяется с подвоем вприклад (камбий на кам бий, корой на камбий) или под кору. При этом важно совпадение диаметров привоя и подвоя.
Окулировка, наиболее распространенный спо соб прививки плодовых, заключается в приращи вании к подвою не черенка, а почки с кусочком коры и древесины (прививка глазком). В коре подвоя делают Т-образный разрез, края коры от гибают от древесины и за кору вставляют глазок, прижимают отворотами и обвязывают. Произво дят прививку летом и весной. В месте прививки образуется каллюс и формируются общие прово дящие элементы, однако строгого расположения их, как в нормальном стебле, не наблюдается. Результат прививки зависит как от техники про изводства прививки, так и от природы компонен тов. Тканевая несовместимость, обусловленная специфичностью белков, слабо выражена между сортами, менее успешны прививки одного вида на другой, и еще труднее — растения одного рода на растение другого рода. Не удаются вообще прививки между одно- и двудольными расте ниями.
При прививках в качестве подвоя используют дички, выращенные из семян сортов или диких видов, имеющих большую устойчивость. Значи тельная работа проведена по выявлению взаимо влияния привоя и подвоя, так как в результате прививки не удается получить растение, полно стью идентичное привою. Для ослабления влия ния привоя рекомендуется черенки и глазки брать со зрелых растений и прививать на моло дые сеянцы-дички.
Для решения многих практических задач в растениеводстве, генетике и селекции сельскохо зяйственных и лесных растений в последнее вре мя большое значение приобретает использование методов культуры тканей и клеток. Из одной клетки, суспензии клеток или группы клеток ткани в асептических условиях выращивают це лые растения для клонального размножения цен ных культур, получения безвирусных форм сель-
261