Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfстолбныё) стебли: все проводящие пучки коллате ральные и каждый из них окружен перициклом и эндодермой. Такую структуру имеют некоторые двудольные (виды лютика, кувшинки). К астелическим относят также стебли, у которых эндодер ма отсутствует.
В дальнейшем стелярная теория неожиданно претерпела изменения. Оказалось, что разнооб разные типы стелы отражают эволюционный ход развития сосудистых растений. На основании этого учения были даны морфологическая харак теристика и филогенетическая оценка многих современных и ископаемых папоротниковидных
иголосеменных растений, особенно псилофитов
иблизких к ним форм. Были установлены типы стелы, присущие крупным таксонам высших рас тений, а главное — эволюционная преемствен ность этих типов. К настоящему времени создана целая номенклатура всевозможных типов стелы, существует модель общей их классификации.
Однако единой оценки стелярной теории пока нет. Одни авторы считают, что стелярная теория систематизировала типы стел основных групп высших растений, выявила эволюционное един ство и общность их строения. Отмечают также, что теория стелы, излагающая главные направле ния эволюции анатомической структуры осевых органов, согласуется с положением теломной тео рии, которая рассматривает эволюционное ста новление побега и корня высших растений. Дру гие авторы полагают, что нет единого взгляда ни на понимание отдельных структур стелы, ни на ее классификацию.
Наиболее примитивным и древним типом сте
лы является протостела (греч. protos — первый и стела). Проводящая ткань образует сплошной тяж ксилемы, окруженный флоэмой (рис. 176, I). Это исходный тип, от которого произошли все
Рис. 176. Схема эволюции стелы:
1 — протостела; 2 — акганостела; 3 — плеетостела; 4 — эктофлойная сифоностела; S — артростела; 6 — амфифлойная сифоностела; 7 — диктяостела; 8 — эвстсла; 9 — атактостела
другие типы стелы. Протостела характерна для древнейших типов наземных растений — риниофитов, плаунов, девонских папоротников. Осевые органы у них еще не были дифференцированы на стебли и корни. Нужно отметить, что и многие современные папоротники на ранних этапах он тогенеза имеют протостелу. Она характерна для плаунов, а также для некоторых водных покрыто семенных. Большинство морфологов-филогени- стов рассматривают центральный цилиндр корня, лишенный сердцевины, как протостелу.
На поперечном сечении протостела может иметь разную форму. Наиболее примитивным ее типом является стела, состоящая из тяжа первич ной ксилемы, окруженной цилиндром флоэмы. Такую протостелу называют гаплостелой (греч. haploos — одиночный, простой). Более специали зированным типом является протостела с ради альными выступами ксилемы, называемая актиностелой (греч. aktis — луч) (рис. 176, 2). Она бы ла характерна для девонского рода астероксшон (Asteroxylon) и встречается у некоторых прими тивных видов плауна. Причины перехода от гаплостелы к актиностеле связаны, во-первых, с по явлением пучков, идущих в боковые органы по бега, и, во-вторых, в актиностеле ксилема имей большую соприкасающуюся поверхность, что спо собствует более активному проведению веществ.
Вопрос, почему у первых наземных высших растений ксилема занимает центральное положе ние, а флоэма — ближе к периферии, до сих пор дискуссионный. Предполагают, что эту черту ор ганизации тела наземные растения унаследовали от водных водорослеподобных предков. Можно допустить, что у водорослей, обитающих в полосе прибоя, механические элементы (прозенхимные, толстостенные клетки) в погруженном слоевище возникли в осевой части. Это обусловило проч ность и гибкость тела водорослей. У сухопутных растений механические клетки приняли на себя функцию водоснабжения и дифференцировались в трахеиды.
Результат дальнейшей специализации актиностелы — появление плектостелы (греч. plectos - плетеный), характерной для стеблей большинства видов плауна (рис. 176, 3). В плектостеле ксилема представлена отдельными участками, располо женными параллельно или радиально друг к дру гу. Как видно, усложнение стелы развивалось в направлении расчленения ее на отдельные тяжи, что выравнивало несоответствие между сильно возрастающим объемом ксилемы и флоэмы и не значительно увеличивающейся поверхностью их соприкосновения с другими тканями. Эти осо бенности отразились в строении и актиностелы, и плектостелы, возникших у растений, для кото рых характерно ветвление телома.
В процессе эволюции протостела дала начало сифоностеле (греч. siphon — трубка, насос). Си фоностела имеет трубчатое строение и обладает центральной паренхимной частью, или сердцеви ной (флоэма, внутри которой расположена кси лема, «охватывает* сердцевину). Возникновение сифоностелы обеспечило существование более крупных растений, так как расположение ксиле-
212
мы на периферии и трубчатый скелет проводя щих элементов сделали стебли более прочными. j Возникновение сердцевины увеличило поверх-
I ность |
проводящих тканей, соприкасающихся с |
||||
|
живыми функционирующими клетками, а также |
||||
|
создало возможность запасания питательных ве |
||||
|
ществ. С появлением растений с крупными ли |
||||
|
стьями (папоротники) формируется сифоностела, |
||||
|
в которой уже есть сердцевина и заметны листо |
||||
|
вые следы и листовые щели (лакуны) — в самом |
||||
|
проводящем цилиндре. Благодаря лакунам созда |
||||
|
ется непрерывная связь коры и сердцевины. Раз |
||||
|
новидности сифоностелы: эктофлойная — ксиле- |
||||
[ ма снаружи обрамлена флоэмой, |
перициклом и |
||||
|
эндодермой (рис. 176, 4); амфифлойная (солено- |
||||
|
стела) — образуются наружный и внутренний |
||||
I |
слои |
флоэмы, перицикла и эндодермы (рис. |
|||
|
176, 6). Наиболее высокоспециализированная |
||||
I |
сифоностела — диктиостела (греч. dictyon — |
||||
I |
сеть; рис. 176, 7). Ее возникновение связано с |
||||
I |
рассечением |
центрального цилиндра листовыми |
|||
I и веточными |
лакунами, заполненными |
парен |
|||
|
химой, |
на отдельные пучки, |
которые |
сохра |
|
|
няют общее расположение по цилиндрической |
||||
|
поверхности. Лакуны, идущие от сердцевины к |
||||
|
коре в радиальном направлении, представляют |
||||
первичные сердцевинные лучи. Различные типы сифоностелы характерны для многих папорот ников.
Особым типом стелы, производным от сифо- i ностелы, является полициклическая сифоностела. j Она формируется у ряда папоротников (орляк, мараттия), когда с возрастом внутри одной стелы образуется вторая, затем третья и т. д. (поли-
циклия).
Самый совершенный вариант эволюционного развития сифоностелы — эвстела (греч. ей — хо роший, истинный). Для нее характерна система анастомозирующих в узлах коллатеральных или биколлатералыгых пучков, слабая разграничен ность листовых щелей и межпучковых зон. Эв-
U стела широко представлена у голосеменных и по крытосеменных двудольных (рис. 176, 8). По од ним данным, эвстела возникла при образовании из протостелы коллатерального пучка (полистелия); по другим — из эктофлойной сифоностелы, расчлененной листовидными прорывами и серд цевинными лучами на отдельные коллатеральные пучки.
Разновидность эвстелы — артростела (греч. irthron — сустав, член) представлена закрытыми пучками, расположенными в паренхиме вокруг центральной полости стелы и соединяющимися между собой в узлах. Характерна для хвощей (рис. 176, 5).
Завершающее звено в эволюции стелы — атактостела (греч. ataktos — беспорядочный). Ей присущи закрытые коллатеральные пучки, слож ная картина прохождения пучков по стеблю (ду гообразные индивидуальные листовые следы) с их разбросанным расположением по всему попе речному сечению стебля (рис. 176, 9). Атактосте ла развилась в результате крайнего рассечения эвстелы и в эволюционном отношении представ ляет собой крайне специализированную форму
центрального цилиндра, характерного в основном для однодольных.
Некоторые авторы выделяют еще стелу корня, которая возникла из гаплостелы, но эволюционно развивалась в ином направлении. Особенности корневой стелы объясняются приспособлением к почвенному питанию, тогда как актиностела по бега дифференцировалась при формировании фотосинтезирующего листостебельного органа.
ВТОРИЧНОЕ АНАТОМИЧЕСКОЕ СТРОЕНИЕ СТЕБЛЯ
У большинства однодольных и некоторых тра вянистых двудольных (например, у лютика) после завершения дифференциации первичных тканей рост стебля прекращается. У основной группы двудольных растений (травянистых и древесных) и голосеменных стебли продолжают увеличивать ся в диаметре даже в участках, завершивших уд линение. Это нарастание в толщину (обхват) на зывается вторичным ростом.
Переход ко вторичной структуре стебля. Вто ричные изменения связаны с деятельностью двух латеральных меристем — камбия и пробкового камбия.
Подробная характеристика камбиальных кле ток изложена в теме «Проводящие ткани». Диф ференциация камбия происходит в зависимости от вида растения по одному из следующих путей.
1.Из сплошного кольца прокамбия возникает непрерывное кольцо камбия, который откладыва ет слои вторичных проводящих тканей.
2.В первичных пучках вначале дифференци руется пучковый камбий, затем между пучками появляются перемычки межлучкового камбия, постепенно сливающегося с пучковым. Образую щийся непрерывный камбиальный слой отклады вает кольца вторичных проводящих тканей.
3.В первичных пучках возникает пучковый и межпучковый камбий, который образует не про водящие элементы, а паренхимные клетки. В ре зультате между пучками формируются либо тон костенная паренхима, соединяющая кору и серд цевину, либо механические элементы, которые вместе с вкрапленными в них пучками образуют прочную трубчатую конструкцию.
4.Межпучковый камбий вообще не образует ся, и проводящие пучки окружены основной па ренхимой. Существует также ряд промежуточных типов.
Формирующаяся вторичная структура стебля определяется способом заложения камбия и дру гими причинами. Многолетние, длительно на растающие в толщину стебли кустарников и де ревьев чаще развиваются по первому или второму типу; недолговечные стебли с ограниченным утолщением чаще сохраняют пучковое строение (типы 3, 4). На заложение и работу камбия боль шое влияние оказывают число и размеры листьев, их расположение на побеге. Наибольшая актив ность камбия отмечена в синтетических пучках.
Ранее говорилось, что камбий образует две формы клеток: удлиненные в вертикальном на-
213

правлении веретеновидные инициалы и вытянутые
вгоризонтальном направлении или отчасти квад ратные лучевые инициали. Вторичная ксилема {древесина) и флоэма (луб) формируются в резуль тате периклинальных делений камбиальных инициалей и их производных. Клетка, откладываемая
всторону поверхности стебля, со временем ста новится флоэмной, а оказывающаяся ближе к сердцевине, — ксилемной. Таким образом, воз никает длинный непрерывный ряд клеток, тяну щийся радиально от камбия: наружу к флоэме и внутрь к ксилеме. Более молодые элементы дре весины образуются дальше от центра стебля, накладываясь на появившиеся ранее, а молодые элементы луба, наоборот, подкладываются под образовавшиеся прежде.
Образованные веретеновидными инициалями клетки ксилемы и флоэмы с их вертикально ори ентированными продольными осями составляют осевую систему вторичных проводящих тканей. Лучевые инициали дают ориентированные гори зонтально лучевые клетки радиальных лучей, или
радиальной системы (рис. 177). Эти лучи состоят из паренхимных клеток и неодинаковы по длине. Питательные вещества переносятся от протопла ста к протопласту по плазмодесмам (симпластный транспорт), проходя от вторичной флоэмы через камбий и далее по радиальным лучам к живым клеткам вторичной ксилемы. В то же время во
да проходит от вторичной ксилемы к камбию и вторичной флоэме главным образом по обо лочкам клеток лучей и осевой системы (апопластный транспорт). Лучи служат также местом запаса.
По мере утолщения стебля объем древесины увеличивается, и камбиальный слой растягивает ся на ее поверхности. Поэтому время от времени инициальные клетки камбия делятся радиально, увеличивая общую поверхность камбиального слоя. При этом число веретеновидных и лучевых инициалей (и новых лучей) увеличивается так, что соотношение между ними во вторичных про водящих тканях поддерживается примерно на од ном уровне. Камбиальные инициали и их недиф ференцированные производные составляют кам биальную зону.
Характер работы камбия выражается в том, что элементов древесины откладывается в 4—5 раз больше, чем элементов луба. Элементы древе сины полностью сохраняются, и их общая масса накапливается из года в год. Элементы луба (коры) в связи с деятельностью феллогена время от времени дифференцируются в корку, которая постепенно слущивается. Поэтому основная мас са многолетнего стебля представлена древеси ной.
Вумеренных зонах камбий зимой находится в состоянии покоя, а весной возобновляет свою деятельность, откладывая за вегетационный пе риод новые слои вторичных проводящих тканей. Сигналом к реактивации камбия служит распус кание почек и возобновление их роста. По-ви димому, этот процесс стимулируется гормоном ауксином, образующимся в развивающихся побе гах и передвигающимся вниз по стеблю. Реакти вации и поддержанию деятельности камбия спо собствуют и другие факторы.
Сезонный ритм работы камбия выражается в образовании зон древесины — годичных колец, хорошо выраженных у большинства деревьев се зонного климата.
Вторичное строение стебля двудольных травя нистых растений. Вторичные изменения в стебле двудольного растения связаны с деятельностью камбия. Началом этих изменений у стебля пучко вой структуры является заложение межпучкового камбия (рис. 178). Он образуется, как уже гово рилось, в сердцевинном луче путем деления кле ток основной паренхимы правильными парал лельными перегородками. Образование камбия начинается из клеток, прилегающих к пучку, за тем распространяется в глубь сердцевинного лу ча. Постепенно межпучковый и пучковый кам бий образуют в стебле сплошное камбиальное кольцо.
Впучке камбий функционирует обычным об разом: внутрь от себя образует вторичную древе сину, кнаружи — вторичную флоэму. Вторичные проводящие ткани оттесняют первичные элемен-
|
Рис 178. Схема перехода от первичного к вторичному строе |
Рис. 177. Схема связи камбия с происходящими из него тка |
нию стебля двудольного растения: |
А — первичное строение; Б — появление кольца камбия; В — развитие |
|
нями — вторичной флоэмой и ксилемой: |
вторичного строения; 1 — эпидермис; 2 — кора; 3 — первичная флоэма; 4- |
1 — веретеновидная иншщаль; 2 — лучевая инициаль; 3 — луч; 4 — вторич |
вторичная флоэма; 5 — пучковый камбий; 6 — межпучковый камбий; 7 - |
ная ксилема; 5 — вторичная флоэма |
вторичная ксилема; 8 — первичная ксилема; 9 — сердцевина |
214

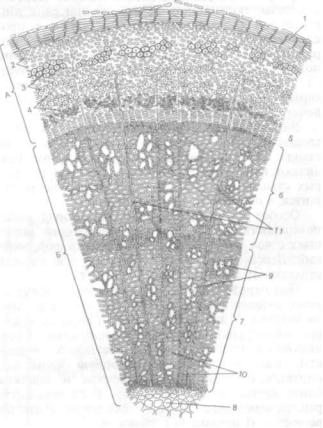
Рис. 179. Стебель клевера ползучего (Trifolium repens) пучкового строения (по перечный срез):
1 — эпидермис с кутикулой; 2 — коровая паренхима; 3 — флоэма; 4 — камбий; 5 — вторичная ксилема; 6 — сердцевина; 7 — сердцевинные лучи с одревес невшими отдельными клетками
ты к периферии пучка, который разрастается и увеличивается в размерах.
Межпучковый камбий формирует новые от крытые проводящие пучки, которые заметно от личаются от пучков прокамбиалъного происхож дения. Они небольшого размера, без механиче ских элементов над пучками. Количество пучков увеличивается, они разрастаются, сливаются ме жду собой, образуя сплошное кольцо вторичной древесины. Внутрь от кольца отдельными высту пами отходят участки первичной ксилемы. Пер вичные сердцевинные лучи при вторичном строе нии суживаются, «сжимаются» древесиной. В свя зи с тем, что в сторону от коры камбий отклады вает меньше клеток, чем в сторону древесины, кора растет медленнее и сохраняет пучковый ха рактер. Такое строение наблюдается в нижних междоузлиях однолетних растений.
Рассмотрим некоторые структурные типы стеб ля травянистого двудольного растения на кон кретных примерах. На поперечном срезе стебля клевера ползучего (рис. 179) видно, что стенки кле ток эпидермиса утолщены и покрыты кутикулой. Под покровной тканью лежит коровая паренхи ма, тонкостенные клетки которой расплющены в тангенциальном направлении вследствие роста стебля в толщину. На более поздних этапах клет ки коровой паренхимы, расположенные непосред ственно под эпидермисом, утолщаются и пре вращаются в колленхиму. У многих травянистых растений с тонкими стеблями имеется пластинча тая или уголковая колленхима, или оба ее типа, что придает стеблям эластичность, упругость и помогает противостоять сильным ветрам, от ко торых ломаются стволы даже крупных деревьев.
Для большинства травянистых двудольных ха рактерно более сильное развитие центрального цилиндра, чем коры. Это хорошо видно у стебля клевера. Крахмалоносное влагалище отсутствует, не дифференцируется и перицикл. Проводящие пучки имеют механическую обкладку.
Центральный цилиндр состоит из проводящих пучков и мощно развитой крупноклеточной серд цевинной паренхимы. Хорошо развит пучковый
камбий, межпучкового пока нет. Проводящие ткани, сформированные пучковым камбием, представлены трахеальными и ситовидными эле ментами. Проводящие пучки соединены в сплошное кольцо межпучковыми секторами ме ханической ткани. Позже над этими секторами возникает межпучковый камбий.
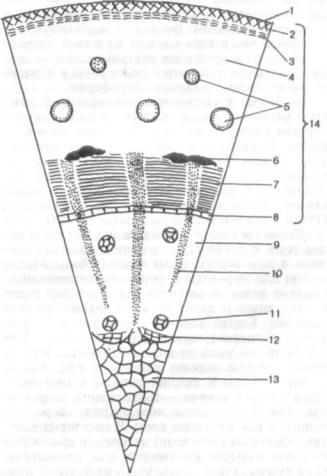
Стебель пучкового строения с хорошо разви тыми сердцевинными лучами и сплошным коль цом механической ткани на границе первичной коры и центрального цилиндра имеет, например, кирказон. Межпучковый камбий закладывается рано, в паренхиме сердцевинных лучей, в связи с чем камбиальное кольцо начинает функциониро вать также сравнительно рано. На рис. 180 пока зано уже вторичное строение стебля кирказона, но с ясно сохранившейся первичной структурой.
Рис. 180. Стебель кирказона (Aristolochia clematitis) с рано возникающим межпучковым камбием (поперечный срез):
1 — кутикула; 2 — эпидермис; 3 — коровая паренхима; 4 — перициклические лубяные волокна; 5 — флоэмная часть пучка; 6 — камбий; 7 — форми рующийся межпучковый камбий; 8 — ксилемная часть пучка; 9 — сердцеви на
215