

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfтканями материнского корня (рис. 142, Б). На кончике клубенька сохраняется апикальная мери стема, благодаря которой он способен нарастать в длину.
Клубеньковые бактерии обладают очень цен ным свойством — фиксировать атмосферный азот, недоступный для непосредственного усвое ния зеленым растением, и превращать его в азо тистые соединения, которые растения усваивают. Клубеньковые бактерии вырабатывают энзимы — катализаторы, которые окисляют свободный азот с помощью кислорода в азотную кислоту. Бобо вые играют важную роль в обогащении почвы азотистыми веществами. Многие из них (клевер, люпин, люцерна и др.) используют в сельском хозяйстве в качестве зеленого удобрения. Эти растения называют сидератами. Так, клевер нака пливает ежегодно до 150—160 кг азота на гектар, люцерна — около 300 кг. В данном случае можно говорить о симбиозе: бактерии питаются за счет растения, а взамен дают азотистые вещества.
При недостатке в почве микроэлемента бора характер отношений меняется: клубеньковые бак терии переходят к паразитизму, превращая расте ние в свою жертву. С другой стороны, в период цветения и плодоношения бобовое растение мо жет «съедать» клубеньковые бактерии, используя их для формирования плодов и семян. В отсутст вие бобовых растений клубеньковые бактерии могут жить в почве как сапрофиты.
Для обозначения вида клубеньковых бактерий принято к родовому названию Rhizobium добав лять название того вида растения, на котором они могут образовывать клубеньки (например, R. trifolii — у клевера, R. lupini — у люпина и т. д.). Активные штаммы клубеньковых бактерий ис пользуют для производства бактериальных удоб рений, например нитрагина.
В последнее время образование клубеньков обнаружено у небобового растения из рода трема (Trema) сем. ильмовых. Некоторые небобовые растения, например лох серебристый, облепиха, также способны фиксировать атмосферный азот. Однако в качестве симбионта служат не бакте рии, а гифы актиномицетов.
Втягивающие корни. Встречаются у растений, обитающих чаще всего в экстремальных условиях
(регионы с сухим жарким климатом или суровой зимой). Такой тип корней, которьш называют еще контрактильные (сокращающиеся) корни, ха рактерен для многих корневищных, луковичных
иклубнелуковичных растений (рис. 143). Как выяснилось, контрактильные корни характерны
идля многих сельскохозяйственных растений (клевер, гречиха, люцерна, морковь, свекла, женьшень). Кроме обычных корней, у этих рас тений развиваются более длинные контрактиль ные корни, которые при сокращении втягивают в почву клубень, луковицу, корневище или гипокотиль и базальную часть побегов с почками возоб новления, т. е. помогают побегам находить опти мальную глубину залегания в почве. При этом контрактильные корни становятся поперечно морщинистыми, благодаря чему их легко отли чить от обычных корней. Это связано с разруше нием участков паренхимы и слабым одревеснени ем тканей. У контрактильных корней, которые выполняют одновременно и функцию запасаю щих органов (например, у спаржи), паренхима не разрушается и поперечные кольцевые морщины не наблюдаются.
Корни-гаустории (лат. haustor — черпающий, пьющий), или присоски. Характерны для парази тирующих растений, таких как омела, повилика, заразиха и др. У омелы, например, паразитирую щей на стволах и ветвях различных древесных растений, образуются длинные цилиндрические или слегка сплюснутые, содержащие хлорофилл корни, растущие в коре растения-хозяина. От них в проводящую ткань отходят присоски, которые поглощают питательные вещества растенияхозяина. Из придаточных почек на этих корнях образуются побеги, пробивающиеся через кору наружу (рис. 144).
Рис. 143. Втягивающие корни (I):
а — крокуса (Crocus vernus); б — кислицы (Oralis acetosella); в — лилии (Lilim martagon)
Рис. 144. Омела (Viscum album) на ветке тополя:
1 — корень-гаустория; 2 — корень в коре тополя; 3 — зачаток побега, разви вающийся из придаточной почки корня
Воздушные корни. Объединяют группу корней, разнообразных по морфологии и часто выпол няющих особые функции. Ходульные корни обра зуются у растений своеобразных растительных сообществ — мангровых зарослей, обитающих по берегам тропических морей в зоне приливов и отливов (рис. 145, А). Ходульные корни — общее популярное название причудливых корней мангров, хотя у разных видов они морфологически разнообразны: «спаржевидные» у авицении из сем. вербеновых; «лентовидные» у ксилокарпуса
182

Рис. 145. Воздушные корни с особыми функциями:
Ходульные корни: А — ризофоры (Rhizophora mangle), образующие мангровые заросли близ устья Риу-Гуаяс в Эквадоре; Б — пандануса (Pandanus); В — кукурузы (Zea mays); Г— досковидные корни у основания ствола дерева в дождевом тропическом лесу, Д— пнематофоры у соннератии (Sonneratia sp.); Е — растения-эпифиты на ветвях сейбы (Ceiba); Ж — часть поперечного среза через воздушный корень орхидеи дендробиум (Dendrobium): 1 — веламен; 2 — пропускная клетка; 3 — экзодерма; 4 — кроющая клетка
гранатового из сем. мелиевых; «ходульные» у ви дов рода ризофора из сем. ризофоровых. Уста новлено, что это дыхательные корни, поглощаю щие кислород во время отливов. Кроме того, об разовавшиеся на стволах мангровых деревьев воздушные корни растут вниз и, укрепившись в почве, прочно удерживают растение в мягком иле.
Ходульные корни часто встречаются у некото рых небольших деревьев тропического дождевого леса, особенно на болотистой почве. Они имеют ся также у некоторых пальм, панданусов (рис. 145, Б), некоторых трав тропического дождевого леса и даже у кукурузы (рис. 145, Б). Способность этих корней-якорей противостоять нагрузкам как на разрыв, так и на сжатие объясняется наличием механических тканей в пучках, расположенных на периферии и в центре корня.
Но наиболее эффектны ходульные корни не которых видов фикуса, особенно фикуса бенгаль ского (Ficus bengalensis), образующего экзотиче скую жизненную форму — баньян. На крупных горизонтальных ветвях взрослого дерева фикуса образуются воздушные корни, гирляндами сви сающие с ветвей. Они растут очень медленно, большая часть их засыхает. Однако физиологиче ское значение этих корней, существенно: в них происходит дополнительный синтез аминокислот в период усиленного роста дерева (А. Л. Курсанов, 1955). Единичные воздушные корни дора стают до земли, укореняются и интенсивно утол
щаются, приобретая внешний облик ствола фи куса и его проводящую функцию. Анатомическое строение такого корневого ствола (корняподпорки) становится близким к строению стеб ля. Благодаря корням-подпоркам крона фикуса разрастается, покрывая площадь до 500 м2.
Досковидные опорные корни наиболее характер ны для крупных деревьев тропического дождевого леса (рис. 145, Г), особенно для деревьев самого высокого яруса. В отличие от ходульных доско видные корни — это боковые корни. Располо женные у самой поверхности почвы или над ней, они развивают более или менее плоские приле гающие к стволу вертикальные надземные вырос ты. В тропическом дождевом лесу досковидные корни нередко поднимаются на высоту человече ского роста. У большинства деревьев с досковидными корнями нет стержневого корня и корневая система состоит из поверхностных боковых кор ней с растущими вниз небольшими ответвления ми. Достаточно удовлетворительной теории, объ ясняющей биологическое значение досковидных корней для деревьев тропического дождевого ле са, в настоящее время нет. Иногда слабо выра женные досковидные корни встречаются и у не которых деревьев умеренной зоны, например у бука, вяза и тополя. В тропическом лесу встреча ются образования промежуточного типа между ходульными и досковидными корнями, а в неко торых случаях у одного растения могут быть и те, и другие.
183
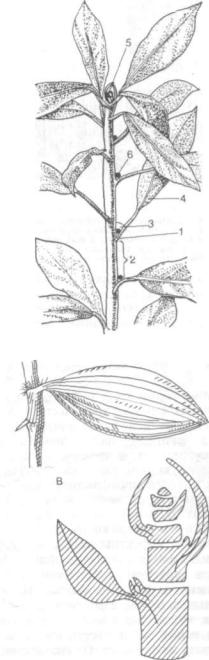
Вентиляционные (дыхательные) корни, или пневматофоры, развиваются у ряда тропических деревьев, растущих на бедных кислородом почвах (рис. 145, Д). Они имеются у мангровых деревьев (например, у авицении), а также у деревьев, про израстающих в лесах на пресноводных тропиче ских болотах. Дыхательные корни образуются из подземных боковых корней и растут вертикально вверх, поднимаясь над почвой или водой, т. е. для них характерен отрицательный геотропизм. Значение пневматофоров заключается прежде всего в снабжении подземных частей корня воз духом, чему способствуют постоянно слущивающаяся тонкая кора, многочисленные чечевички и сильно развитая система воздухоносных меж клетников. Близ поверхности почвы на дыхатель ных корнях образуются тонкие корешки, которые составляют главную всасывающую силу корневой системы мангровых растений. При каждом по следовательном повышении уровня почвы на ды хательных корнях образуются новые группы ко решков.
Воздушные корни тропических эпифитов об разуются у орхидных, ароидных и бромелиевых. Эпифиты (греч. epi — на , над и phyton — расте ние) — растения, поселяющиеся на других расте ниях, главным образом на их стволах и ветвях (рис. 145, Е). Они не паразитируют на деревьях, а используют их в качестве субстрата — опоры для подъема в условия лучшего освещения. Воздуш ные корни эпифитных растений свободно висят в воздухе и приспособлены к поглощению капелек дождя или росы. У них образуется особая много слойная всасывающая ткань — веламен (по про исхождению соответствует ризодермису). Клетки веламена отмирают, и поэтому вода всасывается не осмотическим, а капиллярным путем, прони кая внутрь через большие поры и сквозные от верстия. Изнутри веламен подстилается экзодер мой, через пропускные клетки которой вода пе редается внутрь корня. Пропускные клетки снаружи прикрыты особыми кроющими клетками веламена.
ПОБЕГ
Побег (cormus), как и корень, — основной осевой орган высшего растения. В теле растения побеги бывают вегетативные и репродуктивные.
дают единой проводящей системой. В отличие от корня побег имеет более сложное строение и на ранних этапах онтогенеза расчленяется на сте бель, листья и почки. В целом вегетативный по бег вьшолняет функцию воздушного питания, в то же время каждая его часть (стебель, листья, поч ки) структурно и функционально специализиро ваны.
Стебель представляет несущую основу побега, его осевую часть и вьшолняет две главные функ ции — опорную и проводящую, а иногда — запа сающую. Листья — в типичном случае плоские боковые органы, сидящие на оси, осуществляют главную функцию побега — фотосинтез. Поч-
А
ОБЩАЯ ХАРАКТЕРИСТИКА
Вегетативный побег состоит из стебля — оси побега — и отходящих от него листьев и почек. Репродуктивный побег несет органы размноже ния — спорангии, цветки. Система побегов вме сте с корневой системой составляет тело плауновидных, хвощевидных, папоротниковидных и се менных растений (рис. 146).
В современной ботанике побег трактуется как единый орган того же ранга, что и корень: все час ти побега формируются из общего массива апи кальной меристемы (конуса нарастания) и обла
Рис. 146. Побег:
А — часть побега ивы (Salix): 1 — узел (открытый); 2 — междоузлие; 3 — пазуха листа; 4 — кроющий лист; 5 — верхушечная почка; 6 — боковые (пазушные) почки; Б — закрытый узел побега традесканции (Tradescantia); В — общая схема развития метамеров побега
184
ки — обязательный элемент побега. Как и листья, они формируются в определенном порядке на оси, представляют зачатки новых побегов и обес печивают длительное нарастание побега и его ветвление. В результате деятельности почек обра зуется характерная для вида система побегов. Все три части побега взаимосвязаны происхождением и представляют единый по морфоанатомической структуре орган. Ни один из элементов побега ни образоваться, ни функционировать в отрыве от других не может. Убедительное доказательство единства побега и взаимосвязи всех его частей — явление метаморфоза.
Видоизменение побега в ходе эволюции про исходит в целом. Например, при образовании клубня у картофеля видоизменяется весь побег: стебель разрастается в толщину, в нем преобла дают паренхимные ткани, листья редуцируются, превращаясь в «бровки»; почки («глазки») сохра няют нормальное строение и расположение (см. рис. 229, А).
Структурный элемент побега — метамер (греч. meta — между, через и meros — часть, доля), он представляет узел и нижележащее междоуз лие, а также почку, расположенную в узле (см. рис. 146).
Узел — участок стебля с отходящим от него листом (или мутовкой листьев). У некоторых хвощей, злаков, гвоздичных узлы утолщены и резко выделяются на стебле, у других — границы узлов более или менее условны. Узел может быть открытым, если отходящий лист не охватывает его целиком, или закрытым, если лист или му товка листьев своим основанием полностью ок ружает стебель (рис. 146, А, Б). Участки между соседними узлами называются междоузлиями. На побеге обычно расположено несколько, иногда много узлов и междоузлий, повторяющихся вдоль оси побега, т. е. побег имеет метамерное строе ние.
Главный побег растения, или побег первого по рядка, закладьшается при формировании зароды ша. Он представлен гипокотилем, семядолями, отходящими от семядольного узла, и почечкой, состоящей из стебелька, одного или нескольких листовых зачатков и апикальной меристемы. По чечка считается первой почкой растения, по по ложению — верхушечной, из нее формируются все будущие метамеры главного побега. Верхушечная почка обеспечивает нарастание побега в длину с последовательным формированием новых метамеров.
Вдоль побега, в узлах, располагаются боковые почки. Они прикреплены в пазухах листьев (угол между стеблем и листом) и называются также пазушными почками, лист в таком случае носит название кроющего. Таким образом, каждый ме тамер типичного побега состоит из узла с листом и пазушной почкой и нижележащего междоузлия (рис. 146, В). В меристеме апекса прежде всего вычленяются узлы в виде серии дисков, соответ ствующих будущим листовым зачаткам, а междо узлия разрастаются впоследствии путем интеркалярного роста. Формирующиеся в пазухах листьев боковые почки со временем проходят последова
тельные этапы роста и дифференциации, сходные с наблюдавшимися у первой почки. Эта схема развития в ходе образования побега многократно повторяется. Из боковых почек образуются боко вые побеги, происходит ветвление. Формируется система побегов, состоящая из главного побега и боковых побегов (второго, третьего и других по рядков). Побег любого порядка растет за счет деятельности апикальной меристемы, скрытой в почке на верхушке побега. Следовательно, побег любого порядка в общей системе побегов имеет верхушечную почку.
Система побегов вместе с корневой системой составляет тело типичных высших растений. По беги возникли как приспособление к наземному образу жизни в результате преобразования систе мы безлистных цилиндрических органов — теломов — у риниофитов. Возникновение побегов (т. е. листостебельности) — крупнейший ароморфоз (греч. airo — поднимаю и morphosis — форма, образец; эволюционное преобразование строения и функций организмов, имеющее общее значение для организма в целом и ведущее к морфофизиологическому прогрессу) в истории растительного мира на Земле. Благодаря плоской форме листьев резко возросла фотосинтезирующая поверхность, связанное с этим увеличение транспирации спо собствовало развитию настоящих корней как со вершенных органов поглощения воды и мине ральных солей.
ПОЧКА
Почка (gemma) представляет собой зачаточ ный побег с очень укороченными междоузлиями.
Вегетативная почка состоит из короткой зача точной оси (стебля) с конусом нарастания на верхушке и тесно расположенных на оси разно возрастных зачатков листьев, прикрывающих ось и друг друга, т. е. из серии зачаточных метамеров. В пазухах листовых зачатков почки уже могут быть заложены зачатки пазушных почек следую щего порядка, т. е. уже в почке проявляется по тенциальная способность побега к неограничен ному ветвлению (рис. 147, а).
Генеративная почка содержит зачатки цветков или соцветий (рис. 147, б). Цветочная почка, не сущая цветок, называется бутоном. Бывают веге тативно-генеративные (смешанные) почки, в ко торых заложен ряд вегетативных метамеров и за чаточный цветок или соцветие (рис. 147, в). Такие почки обычны для травянистых (копытень европейский), но бывают и у древесных (сирень, бузина) растений.
Листовые зачатки развиваются в конусе на растания снизу вверх (в акропетальном порядке) и вследствие более ускоренного роста на внеш ней стороне изгибаются к верхушке, образуя замкнутую почку. Наружные листовые органы защищают внутренние части от высыхания и по вреждений и создают внутри почки темную влажную камеру, в которой сохраняется и «ра ботает» меристема.
Наружные листья или их части иногда, видо-
185
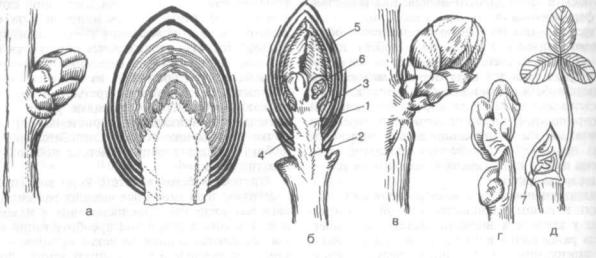
Рис. 147. Типы почек (по А. Е. Васильеву и др., 1978):
а — закрытая вегетативная почка дуба (Quercus robur), общий вил и продольный разрез; б — закрытая генеративная почка вишни (Cerasus vulgaris),продольный разрез; в — закрытая вегетативно-генеративная почка бузины (Sambucus racemosa); г — открытая почка настурции (Nasturtium officinale); д — открытая почка клевера (Trifolium repens); 1 — конус нарастания; 2 — зачаточный стебель; Л — зачаточные листья; 4 — пазушные почки; 5 — почечные чешуи; 6 — зачаток цветка (соцветия); 7 — прилистник
изменяясь, превращаются в почечные (кроющие) чешуи. Почки, имеющие кроющие чешуи, назы ваются закрытыми. Они характерны главным об разом для многолетних древесных растений хо лодного и умеренного поясов — дуб, береза, ли па, вишня, лещина, ольха, бузина, ива И др. (рис. 147, а — в), а также для субтропических и тропи ческих областей с сухим периодом. У многих рас тений почечные чешуи пропитаны густым клей ким слизистым веществом (каштан конский, то поль бальзамический), у железного дерева чешуи почти черные, имеют множество волосков, повидимому, пигментация чешуи играет защитную физиологическую роль. Войлочное опушение имеют почечные чешуи дуба каштанолистного. Число почечных чешуи у разных видов различно: иногда их свыше 20 (дуб), иногда всего две (ива) или даже одна (почки некоторых однодольных). При развертывании закрытых почек чешуи опа дают, оставляя почечные кольца, по которым мож но определить границы побегов разного года и возраст ветви.
Открытые, или голые, почки лишены специ альных видоизмененных чешуи (рис. 147, г, д). Открытые почки на верхушке имеют весной и летом растущие побеги многих деревьев и кус тарников, у которых зимой были закрытые поч ки, а также побеги многих однолетних и много летних трав. Некоторые травянистые растения зимуют с открытыми почками (кошачья лапка, зеленчук, живучка и др.). Открытые почки харак терны для многих древесных пород влажных тро пических лесов. У открытых почек конус нарас тания окружен листовыми зачатками разного воз раста, а у некоторых растений прикрыт прилист ником предыдущего листа (клевер, рис. 147, д) или «своего» листа (береза). У злаков верхушеч ная почка растущего побега заключена в трубку из влагалищ одного или нескольких более взрос лых листьев.
Положение почек на побеге. Как уже известно, почки по положению на побеге бывают верхушеч
ные, или апикальные, и боковые (пазушные). Боль шая часть боковых почек на следующий год по сле их закладки распускается и образует новый побег. Такие почки называются активными. Па зушное положение почки имеет важное биологи ческое значение: кроющий лист защищает почку от высыхания и механических повреждений и интенсивно снабжает ее ассимилятами.
Положение почек в пазухе листа. По положе нию в пазухе листа почки бывают одиночные (вишня, яблоня, каштан конский и др.); сериаль ные, если почки расположены в один вертикаль ный ряд (жимолость татарская, аморфа, кирказон, робиния, грецкий орех); бисериальные, если почки располагаются в два параллельных вертикальных ряда; коллатеральные, или горизонтально-рядовые,
если почки расположены в один горизонтальный ряд (акация, волчье лыко, лук, инжир); биколлатералъные, если почки располагаются в два па раллельных (горизонтальных) ряда; мутовчатые, если почки в пазухе листа располагаются как бы по окружности — мутовками (слива войлочная). Почкорасположение показано на рис. 148.
На месте опавшего кроющего листа на стебле образуется листовой рубец (листовая подушка), на котором хорошо заметны следы проводящих пуч ков, идущих от листа в стебель (листовые следы, рис. 148, 7).. Форма, размеры листовых рубцов и листовых следов специфичны и служат критери ем определения видов в безлистном состоянии.
Спящие почки. По функциональному значе нию выделяют спящие почки, которые не рас пускаются ежегодно и остаются живыми в тече ние многих лет. На стеблях древесных растений они многочисленны, при утолщении ствола спя щие почки ежегодно нарастают, образуя в толще скрытую ветку. Их активное состояние начинает ся после удаления вышерасположенной части стебля. В пазухах чешуи спящей почки заклады ваются дочерние почки, которые могут подрас тать и тоже давать скрытые ветви, т. е. спящая почка ветвится. Благодаря этому у основания
186

Рис. 148. Почхорасположение:
1 — очередное у вяза (Ulmus); 2 — супротивное у клена остролистного (Acer platanoides); 3 — сериальное у сифизии (Arislolochia sipho); 4 — бисериальное у кирказона обыкновенного (Aristolochia clematitis); 5 — коллатеральное у волчьего лыка (Daphne mezereum); 6 — мутовчатое у войлочной сливы (Prunus tomentosa); a — верхушечная почка; 6 — пазушные; 7 — листовые рубцы с листовыми следами (s) соответственно у дуба длинноножкового (Quercus longipes), айланта высочайшего (Ailanthus auissima), сливы домашней (Prunus domestica)
стволов их накапливается большое количество (рис. 149).
В практике озеленения и садоводства разрабо таны специальные приемы обрезки, учитываю щие местоположение и физиологическую осо бенность спящих почек. Таким путем стимулиру ется развитие кроны у плодовых деревьев, при дается желательная форма кроне декоративных древесных растений, омолаживаются старые дере вья. В естественных условиях из спящих почек образуется корневая и пневая поросль после от мирания или рубки дерева, так происходит во зобновление леса. Спящие почки формируют систему скелетных осей кустарников (лещина, смородина, жимолость и др.), которые периоди чески отмирают. Побеги, сформированные из спящих почек, на крупных ветвях многих плодо вых деревьев называют волчками (водяными или жировыми побегами). В практике садоводства волчки чаще всего вырезают, так как их более
обводненные ткани медленнее вызревают, а сами волчки загущивают крону.
У некоторых растений спящие почки на ство ле образуют безлистные цветоносные побеги. Та кое явление носит название каулифлории (греч. kaulos — стебель, ствол и лат. flos, род. п. floris — цветок) и характерно для многих деревьев тропи ческих лесов (какао, фикус; рис. 149, 3). У гледи чии, субтропического декоративного дерева сем. бобовых, из спящих почек на стволе вырас тают пучки крупных разветвленных колючек (рис. 149, 4).
Нередко спящие почки не пробуждаются в те чение всей жизни растения и погибают вместе с побегом или корнем. По происхождению спящие почки могут быть пазушными и придаточными.
Придаточные, или адвентивные, почки. В от личие от спящих придаточные почки могут фор мироваться на всех частях и органах растения — стеблях, листьях, корнях. В отличие от пазушных,
Рис. 149. Спящие почки:
1 — у основания ствола березы (Betula verrucosa); 2 — схема на растания спящей почки (про дольный разрез); 3 — каулифло рия у фикуса пестрого (Ficus variegata); A — колючки у гледи чии (Gleditsia triacanthos) из спя щих почек
187
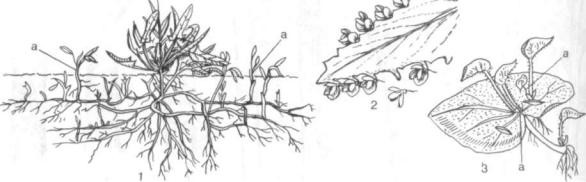
Рис. ISO. Образование побегов (а) из придаточных почек:
/ — у щавелька (Rumex acetosella) на корнях (корневые отпрыски); 2 - из выводковых почек на листьях каланхое Дегремона (Kalanchoe daigremonttana); 3 — на листьях бегонии (Begonia rex)
которые образуются экзогенно в апексе побега, придаточные почки возникают из активизирую щихся паренхимных клеток взрослого органа эн догенно. Источником их образования служат кам бий, перицикл, раневые меристемы, паренхима сердцевинных лучей, мезофилл, эпидермис. Ог ромное количество придаточных почек, обычно образующихся у растений, имеет большое биоло гическое значение, так как они обеспечивают вегетативное размножение и возобновление. У многих растений из придаточных почек на корнях образуются корневые отпрыски (рис. 150, 7). К корнеотпрысковым растениям относятся осина, малина, одуванчик, щавелек, иван-чай, лыгянка обыкновенная, осот желтый и лиловый (бодяк). В естественных условиях корневая и пневая поросль может образовываться и из при даточных почек.
На листьях придаточные почки формируются реже. У многих растений из рода каланхое (виды каланхое, бриофиллюм) сем. толстянковьгх (Crassulaceae) придаточные почки, называемые вывод ковыми, располагаются по краю листа. Прорастая в маленькие побеги с придаточными корешками, они отпадают и вырастают в новые особи (рис. 150, 2). Выводковые почки образуются на листьях сердечника, насекомоядного растения росянки, у многих папоротников.
В результате деятельности придаточных почек легко размножаются листовыми черенками бего ния, герань душистая, ирезина, эхеверия (рис. 150, 3).
Почки возобновления. По происхождению мо гут быть и экзогенными (апикальными или боко выми) и эндогенными (придаточными). За счет деятельности этих почек происходит постоянное нарастание системы побегов. Для них обязателен период покоя, поэтому в сезонном климате с зимним периодом их называют зимующими, в об ластях без зимы — покоящимися. После периода покоя почки возобновления регулярно функцио нируют, обеспечивая многолетнее существование древесного или травянистого растения.
Степень сформированности будущего побега в покоящейся почке возобновления различна.
У одной группы растений умеренного и холодно го климата в зимующих почках к осени сформи рованы полностью и вегетативный, и генератив ный побеги, иногда до зрелой пыльцы в пыльни ках. Сюда относятся все цветущие весной дре весные растения средней полосы, а также раноцветущие травянистые многолетники (печеноч ница, гусиный лук, пролеска, ветреницы, чистяк, медуница и другие «подснежники»), лютики, земляника. У другой группы растений в почках возобновления к осени закладываются вегетатив ные побеги, а весной, после перезимовки, идет развитие цветков и соцветий. Сюда относятся растения, цветущие в середине лета (боль шинство луговых трав, растения опушек, при брежий). У растений третьей группы в зимующих почках возобновления к осени наблюдаются только начальные этапы развития вегетативных органов, окончательное формирование вегетатив ных и генеративных побегов происходит весной и летом в процессе открытого роста побега. Такие растения цветут поздно летом, обильно олиственны — золотая розга, иван-чай, многие астровые. Следовательно, сроки цветения растений нахо дятся в прямой корреляции с динамикой разви тия почек возобновления.
Покой почек. Это одно из проявлений общего покоя растений, обусловленное сезонностью климата. Это особое физиологическое состояние, при котором резко снижаются скорость роста и интенсивность обмена веществ, распускание по чек. Покой почек — приспособление для пере живания неблагоприятных внешних условий в определенные периоды жизненного цикла или сезона года. В период покоя повышается способ ность почек переносить без повреждения недос таток воды, низкие температуры. Различают глу бокий покой, при котором рост почек не возоб новляется даже при оптимальных условиях. Это обусловлено внутренними факторами — повыше нием содержания в тканях ингибиторов роста. Вынужденный покой вызван внешними фактора ми, он прекращается с наступлением благоприят ных условии. Почки древесных растений осенью впадают в глубокий покой, который в конце зи-
188
мы, задолго до их распускания, сменяется выну жденным покоем. Только после воздействия низ ких температур, эволюционно необходимых рас тениям в условиях сезонного климата, почки тро гаются в рост. В тропических и субтропических поясах с выраженным засушливым периодом ве гетация почек прерывается летним покоем.
Почки обогащения. Если пазушные почки со всем не имеют периода покоя и начинают рас пускаться одновременно с ростом основного по бега, их называют почками обогащения. Образую щиеся из них побеги обогащения увеличивают (обогащают) фотосинтетическую поверхность ра стения, число соцветий и, следовательно, семен ную продуктивность. Такие почки и развиваю щиеся из них побеги характерны в основном для большинства однолетних трав (василек синий, лебеда, иван-да-марья), а также для ряда много летних трав с удлиненными цветоносными побе гами (василек луговой, вербейник обыкновен ный). Побеги обогащения образуют одно возраст ную с материнским побегом систему, отмираю щую у трав целиком, одновременно. У древесных растений побеги обогащения встречаются редко, иногда их можно наблюдать на крупных водяных побегах (волчках). Однако у сосны обыкновенной пазушные почки распускаются одновременно с ростом материнского побега, формируя мутовку побегов обогащения — брахибласты.
Развертывание побега из почки. У большинст ва растений средней полосы почки распускаются весной, с наступлением устойчивых положитель ных температур. Начало распускания почек — генетически обусловленный признак каждого таксона, но внешние условия могут несколько изменить эти сроки. Начало превращения почки в побег связано с ростом зачатков листьев и уве личением междоузлий. Листья, шатром укрывав
шие апекс побега, начинают усиленно расти на стороне, обращенной к нему, изгибаются кнару жи, обнажая ось. Наружные чешуи обычно не растут или растут слабо, постепенно засыхают и опадают, оставляя почечные кольца. Внутренние почечные чешуи некоторое время растут основа нием, в связи с чем весной окраска верхних, зи мовавших частей чешуйки бурая, а нижних — светлая, разных тонов — от зеленой до розовой в зависимости от вида растения. Позже эти чешуи тоже опадают. Выход из почки зеленых фотосинтезирующих листьев связан с усиленным разрас танием его частей. У зачаточного стебля начина ют удлиняться междоузлия в результате деятель ности вставочной меристемы, сохраняющейся в узлах. Развернувшийся из почки побег с хорошо выраженными, крупными междоузлиями называ ют удлиненным побегом (у древесных растений та кие побеги обычно называют ростовыми; см. рис. 146, А; 151, А). Если междоузлия растут медленно, образуется укороченный побег. Каждый год такие побеги удлиняются лишь на несколько милли метров и редко ветвятся. Их легко узнать по не большой длине и многочисленным почечным кольцам и листовым рубцам, остающимся от тес но скученных опавших листьев и почечных че шуи. Хорошо выражены укороченные побеги то поля, березы, бука, платана, осины (рис. 151, А, Б). У плодовых (рис. 151, В) укороченные побеги называют «плодушками», только на них образу ются цветки и плоды. Очень характерны укоро ченные побеги, несущие пучки хвои, у некоторых хвойных (сосна, лиственница). Укороченными побегами можно считать и стебли, развивающие розетки листьев, например у первоцветов, оду ванчика, подорожника, моркови, свеклы и других корнеплодов (в первый год жизни), агав, так на зываемые розеточные побеги (рис. 151, Г). Многие
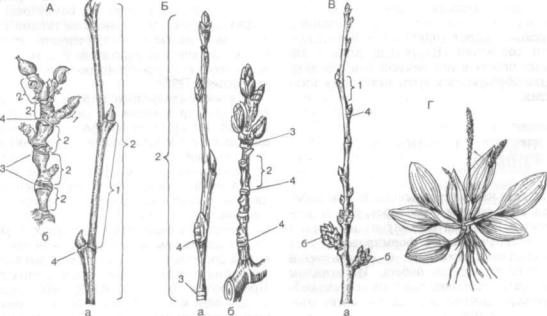
Рис. 151. Удлиненные (а) и укороченные (б) побеги:
А — платана (Platanus orientalis); Б — осины (Populus tremula); S — вишни обыкновенной (Cerasus vulgaris); Г— розеточный побег подорожника (Plantago major) с пазушными цветоносами; 1 — междоузлие; 2 — годичный прирост; 3 — почечные кольца; 4 — лис товой рубец
189
т^Н шш^^шшш^^шш^^^^^^^^шш |
I^HB |
|
укороченные побеги быстро теряют верхушечную почку, замедляют развитие и опадают.
Верхушечная почка ростовых побегов форми рует новые метамеры, т. е. работает как морфоло гически отграниченная его часть, постоянно са мообновляющаяся и восстанавливающаяся. Если верхушечная почка по различным причинам (повреждение, подмерзание) погибает, рост ство ла или крупной ветви дерева может продолжаться за счет деятельности боковых почек (см. Вет вление побега). Верхушечная меристема, форми рующая цветок или соцветие, полностью расхо дуется, и побег больше не способен к верхушеч ному росту.
Развертывание побегов в сезонном климате умеренной зоны у большинства растений носит периодический характер. У древесных растений и многих многолетних трав побеги формируются из почки раз в году — весной или в начале лета, по сле чего закладываются новые зимующие почки с зачатками побегов будущего года. Побеги, вырас тающие из почек за один вегетационный период, раз в год, называют годичными побегами или го дичными приростами. Границами годичных побе гов разных лет являются почечные кольца, ос тавшиеся от развернувшейся верхушечной почки. У листопадных деревьев листья образуются толь ко на годичных побегах текущего года, у вечнозе леных — листья часто сохраняются на годичных приростах 3—5 прошлых лет.
В бессезонном климате влажных тропиков рост многих деревьев тоже строго периодичен. На некоторое время у них образуются покоящиеся почки, потом они развертываются в побеги, фор мируются новые почки и т. д. Это повторяет ся 3—7 раз в году. Ряд субтропических культур (цитрусовые, чайный куст) также дают по 3—4 прироста, разделенных небольшими периодами покоя. Такие побеги, образующиеся за один цикл роста несколько раз в году, И. Г. Серебряков (1952) предложил называть элементарными побе гами. При одном ростовом цикле в сезонном климате средних широт годичный и элементар ный побеги совпадают. В средней полосе два элементарных побега в год (весной и в середине лета) нередко образуются у дуба, некоторых мно голетних трав.
СТРОЕНИЕ И ДЕЯТЕЛЬНОСТЬ АПЕКСА ПОБЕГА
Терминальная часть верхушечной почки побе га представлена апексом (лат. apex, род. п. apicis — вершина), который состоит из первичной меристемы, обеспечивающей формирование всех частей и первичных тканей, т. е. органогенез и гистогенез развивающегося побега. Источником постоянного самовосстановления апекса являют ся инициали верхушечной меристемы, сосредото ченные на кончике апекса.
Как известно, апекс корня всегда гладкий. На апексе побега регулярно формируются выступы (экзогенно) — бугорки или валики зачатков ли
стьев, так называемые листовые примордии (лат. primordialis — первоначальный). Они появляются в акропеталъной последовательности, снизу вверх. Гладким у апекса побега остается лишь кончик, его дисталъная часть (лат. distalis — наиболее уда ленный от центра), которую называют конусом нарастания или точкой роста побега. По форме и размерам апексы могут быть неодинаковыми не только у разных видов, но и у одного растения или отдельного побега в разные периоды его морфогенеза. Средние диаметры апексов на уров не самого молодого листового бугорка — 100500 мкм.
Видимые изменения апекса связаны с после довательным вычленением метамеров, состоящих из междоузлия и узла с пазушной почкой. Зако номерность чередования метамеров от основания к верхушке легко наблюдается в апексе побега и даже еще в зародыше семени. Зачатки листьев - примордии — так плотно сомкнуты друг с дру гом, что формирующийся побег представляется в виде дисков, как бы наложенных друг на друга. Диски — зачаточные узлы будущего побега - в апексах структурно однородны. В дальнейшем они подвергаются значительной морфофизиологической дифференциации, что определяет осо бенность структуры развивающегося побега. Позднее у основания дисков начинает функцио нировать интеркалярная меристема, ее деятель ность определяет длину междоузлий. Развитие дисков-метамеров у разных природных групп происходит по-разному: у одних — с явным пре обладанием осевой, т. е. стеблевой, структуры, у других — листовой. Первая наиболее ярко выра жена у простейших наземных растений — риниофитов и голосеменных, так называемый артрофитный, ярко выраженный членистый тип побе га; вторая — у папоротников — филофитный тип. У покрытосеменных одни группы растений (на пример, банановые) отражают филофитный тип строения, другие (например, бамбуковые) - артрофитный. Между названными типами существу ет целая гамма переходов. Членистая артрофитная структура хорошо выражена у злаков, развитие метамеров которых детально изучено Т. И. Се ребряковой (1978).
Наличие столь ярко выраженных двух основ ных структур побегов — артрофитной и филофитной — привело морфологов к длительной дискуссии о природе побега. Сторонники кауломной (лат. caulis — стебель) теории главным в побе ге считают его ось. Сторонники фолиарной (лат. folium — лист) теории за основу побега признают лист, а стебель считают производным оснований листьев.
Заложение метамеров происходит ритмично. В результате вычленения очередного метамера объем дистальной части апекса уменьшается, а затем опять нарастает до определенных размеров. Промежуток времени между вычленением на апексе двух последовательных метамеров называ ется пластохроном (греч. plastos — форма, вылеп ленный и chronos — время).
Продолжительность пластохрона у различных растений, у одного растения и даже у одного по-
190
г
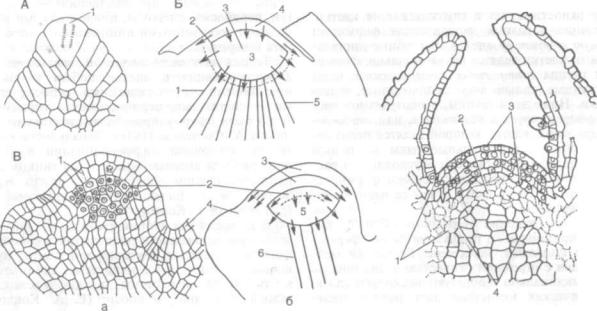
Рис. 152. Строение верхушки побегов (по К. Эсау, 1980):
А — хвоща (Equisetum hyemale): 1 — апикальная клетка; Б — сосны (Pinus sflvestris), схема продольного разреза: 1 — периферическая зона; 2 — переходная зона; 3 — центральные материнские клетки; 4 — инициальный слой; 5 — колончатая меристема; В — гороха (Pisum sativum): a — детальное строение, 6 — схема; 1 — антиклинальное деление; 2 — инициальный слой корпуса; 3 — двухслойная туника; 4 — листовой примордий; 5 — корпус; 6 — сердцевинная меристема; Г — колеуса (Coleus blumei): I — периферическая меристема; 2 — центральные материнские клетки (зона примерно соответствует корпусу); 3 — туника; 4 — сердцевинная меристема
бега в разные периоды его роста может быть не одинакова. В период активного весеннего роста у большинства древесных растений он равен в среднем 2—4 суткам. Чем мельче листовые зачат ки, тем короче пластохроны.
Кроме ритмичных изменений, связанных с вычленением метамеров, апекс побега испытыва ет возрастные, онтогенетические изменения, ко торые накладываются на пластохронные. Резкие изменения, связанные с биохимическими и фи зиологическими процессами, происходят в апексе в связи с наступлением генеративной фазы. Фор ма апекса меняется, и вместо листовых зачатков на нем начинают формироваться зачатки частей верхушечного цветка или отдельных цветков со цветия. Это уже не вегетативный, а флоральный (лат. flos, род. п. floris — цветок) апекс. Следова тельно, апекс побега является тем местом, где происходят важнейшие структурно-физиологи ческие процессы, ведущие к образованию вегета тивных и генеративных органов.
К. Ф. Вольф (1759) впервые установил, что верхушка побега имеет недифференцированное строение и из ее клеток в дальнейшем развивают ся все ткани надземных органов растения. Счита лось, что в верхней части побега расположена апикальная клетка, которая представляет струк турно-функциональную единицу верхушечных меристем, управляющую всем процессом роста.
Теория апикальной клетки применима к большин ству высших споровых растений, у которых на верхушке апекса имеется действительно одна инициальная клетка (рис. 152, А). Эта клетка иногда двугранной, но чаще тетраэдрической формы, крупная и сильно вакуолизированная. Как правило, она делится параллельно своим по верхностям, ее производные составляют осталь ную часть апекса, участвуют в образовании лис-
товых зачатков, затем после ряда делений диффе ренцируются в первичные ткани побега.
Интенсивное исследование апексов побегов и зародышей, проведенное Дж. Ганштейном (1868, 1870), позволило обосновать теорию гистогенов. Ее основные положения: 1) тело растения возни кает не из поверхностных клеток, а из массы ме ристемы, расположенной внутри апекса; 2) весь массив меристемы состоит из трех слоев, или гистогенов, которые отличаются по происхожде нию и развитию. Самый наружный (первый) гистоген — дерматоген — является зародышевой по кровной тканью, второй гистоген — периблема — дает начало коре, а третий — плерома — форми рует внутренние ткани. Дерматоген и периблема образуют слои, которые подобно мантии, покры вают плерому. Дерматоген, каждый слой периб лемы и плеромы возникают из одной или не скольких инициалей, расположенных рядом друг над другом в наиболее дистальной части апи кальной меристемы.
Термин «дерматоген» Дж. Ганштейна не явля ется синонимом термина «протодерма», введенно го Г. Габерландтом (1914). Протодермой называют самый наружный слой апикальной меристемы независимо от того, возникает этот слой из само стоятельных инициалей или нет и дает он начало только эпидермису или и субэпидермальным тка ням. Теория гистогенов была разработана как для апекса корня, так и для апекса побега. Однако в настоящее время она во многом утратила свое значение, и если рассматривается, то примени тельно к апексу корня (см. рис. 132).
Уголосеменных и покрытосеменных растений апекс побега состоит из многоклеточного массива меристемы. Клетки апекса неоднородны по раз мерам, форме, темпу и направлению делений.
Уголосеменных (рис. 152, Б) группа иници-
191