

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfРис. 126. Строение молодого растения конских бобов (Vicia faba). Показаны основные вегетативные органы и ткани выс шего растения
ния боковых частей органа по отношению к оси. В морфологии растений под симметрией пони мают такую структуру, при которой плоскость симметрии, проходящая через ось, рассекает ор ган на две тождественные (зеркально подобные) части. В связи с этим различают три группы ор ганов — несимметричные, моносимметричные (греч. monos — один) и полисимметричные.
Одна из общих морфофизиологаческих зако номерностей вегетативных органов — полярность. Каждый орган имеет два полюса: верхний, или верхушечный, — терминальный (лат. terminalis — верхушечный), апикальный; нижний — базальный (греч. basis — основа, основание).
Физиологические различия между апикальной и базальной частями растений находят свое вы ражение в явлении тропизма (греч. tropos — по ворот). Тропизмы связаны с воздействием света, силы тяжести, химических и других раздражите лей, сообразно которым их называют фототропизмами, геотропизмами, хемотропизмами и т. д. Если изгиб происходит в сторону раздражающего фактора, тропизм положительный, если в проти воположную сторону — отрицательный. Главный стебель обычно растет вертикально и одновре менно тянется к свету, т. е. обладает отрицатель
ным геотропизмом и положительным фототро пизмом. Главный корень всегда растет в землю и избегает света, т. е. обладает положительным гео тропизмом и отрицательным фототропизмом.
Корень и стебель обычно располагаются вер тикально, образуя единую ось, и являются поэто му осевыми органами, несущими на себе боковые придатки (листья, шипы, волоски и т. д.).
Итак, вегетативное тело любого высшего рас тения (кроме моховидных) в каждый момент его жизни состоит из побеговой и корневой систем, которые построены из повторяющихся элементов структуры — побегов и корней. В течение жизни у растения происходит как отмирание старых час тей, так и образование новых (вначале семядолей, листьев, молодых корешков, затем более крупных участков побеговой и корневой систем).
Таким образом, тело растения постоянно ме няет свое положение в пространстве и изменяет свой облик во времени.
В процессе эволюционного развития вегета тивные органы претерпевали видоизменения (ме таморфоз), обусловливая приспособление расте ний к различным условиям обитания.
КОРЕНЬ
Корень (radix) — один из основных вегетатив ных органов листостебельных растений, служа щий в типичном случае для поглощения воды и питательных веществ из почвы. Корень — осевой орган, обладающий, как правило, цилиндриче ской формой, радиальной симметрией и неогра ниченным ростом за счет деятельности апикаль ной меристемы.
Корень морфологически отличается от побега тем, что обладает положительным геотропизмом и гидротропизмом, его апикальная меристема всегда прикрыта корневым чехликом, и на корне нико гда не формируются листья, в связи с чем нет интеркалярного роста. В редких случаях корни образуют придаточные почки, из которых разви ваются придаточные побеги.
В связи с главной функцией всасывания рас творов из почвы для корня характерны интенсив ное ветвление и огромная внешняя поверхность, что позволяет более полно освоить почвенное пространство. Во внутренней структуре корня в связи с этим важное место занимают специальная поглощающая ткань и проводящие ткани.
Поглощая из почвы воду и растворенные в ней вещества, корни обладают способностью по давать эту воду вверх в стебель и листья. Способ ность корней нагнетать в стебли воду из почвы называется корневым давлением, которое у рас тений обычно равно 0,2—0,3 МПа (2—3 атм), у некоторых значительно выше. Растения, обла дающие очень высоким корневым давлением, мо гут расти в сухих степях и пустынях, где они до бывают воду из глубоких слоев почвы. Подача корнем воды вверх по стеблю бывает хорошо за метна весной на пнях, на поверхности которых выделяется большое количество сока.
Корни, как и все другие органы растений,
162
дышат, используя кислород почвы и выделяя в почву углекислый газ. Корни растений менее разнообразны, чем стебли и листья, так как среда их обитания довольно однообразна.
Помимо главной функции — поглощение ве ществ из почвы, корни вьшолняют ряд других физиологических и механических функций:
1)закрепляют («заякоривают») растения в почве, делают возможным вертикальный рост и вьшесение побегов вверх, обладают огромной прочностью на разрыв. Например, чтобы выдер нуть из почвы молодое растение кукурузы, надо приложить силу около 100—130 кг;
2)в корнях происходит синтез различных ве ществ (многих аминокислот, алкалоидов, гормо нов и пр.), поступающих затем в другие органы растений. Многие авторы установили, что роль корня в образовании органических веществ неза менима;
3)служат органом запаса, а нередко преобра зуются в запасающий орган (например, у свеклы, моркови, редьки);
4)взаимосвязаны с другими организмами, на селяющими почву. Своими выделениями корень способствует симбиозу с грибами и почвенными бактериями, которые также участвуют в процессе поглощения и превращения веществ;
5)с помощью корней или их частей растения могут вегетативно размножаться.
Перечисленные функции характерны для большинства нормально развитых корней. У мно гих растений корни вьшолняют особые функции.
Типичные корни, у которых главный корень развивается из зародышевого корешка и форми рует корневую систему, встречаются преимущест венно у голосеменных и покрытосеменных расте ний. Хотя и здесь бывают исключения: отсутствуют корни у некоторых водных растений (вольфия бес корневая, пузырчатка), а также у взрослых парази тических растений (омела, повилика).
ЭВОЛЮЦИОННОЕ ПРОИСХОЖДЕНИЕ КОРНЯ
Филогенетически корень возник позднее, чем стебель, и, вероятно, произошел от корнеподобных образований первых наземных растений — ршшофитов (см. гл. 7). Тело риниофитов еще не было дифференцировано на побеги и корни, а состояло из осевых органов, нараставших в длину за счет верхушечных меристем и дихотомически ветвившихся. Одни из ответвлений располагались вертикально, другие расстилались по земле, обра зуя корнеподобные веточки — ризомоиды, и впи тывали водные растворы минеральных солей. Риниофиты были растениями небольших размеров, обитали у кромки воды и в воде, в местах с влажным климатом. Отсутствие корней и прими тивное строение не мешали их нормальному обеспечению водой. Подобные ризомоиды из ныне живущих растений сохранились только у псилофитовых.
В процессе дальнейшего развития отдельные ризомоиды стали углубляться в почву и дали на
чало корням, приспособленным к более совер шенному почвенному питанию. Появление кор ней как специализированных органов почвенного питания вызвало всеобщую перестройку их структуры, появление специализированных тка ней. Настоящие корни первоначально появились у плауновидных и папоротниковидных. Эволю ционное развитие корня привело постепенно к функциональной дифференциации отдельных его частей: поглощать водные растворы из почвы стали молодые участки корешков, появилась осо бая всасывающая ткань — ризодермис, более ста рые участки корня приняли на себя функции «заякоривания», запаса и т. д. «Оседлый» образ жизни растений требовал освоения больших площадей почвенного питания, что обеспечива лось увеличением общей поглощающей поверх ности корней за счет ряда факторов. Главные из них: а) ветвление и возникновение все большего числа всасывающих окончаний; б) постоянный рост корней и передвижение их в новые участки почвы; в) образование специальных структур вса сывания — корневых волосков; г) образование новых придаточных корней. Возникло новое структурное образование для защиты растущего участка корня — корневой чехлик.
Образование корней было крупным эволюци онным шагом в развитии наземных растений, оно обеспечило завоевание засушливых местообита ний, позволило растениям «подняться» над зем лей, что наряду с другими факторами привело к появлению крупных растительных форм.
Из нынеживущих сосудистых растений корни отсутствуют у наиболее древних и примитивных порядков риниевых (Rhyniales) и псилофитовых (Psilophytales). Нет корней у мхов. Их роль вы полняют ризоиды, отличающиеся от корней от сутствием тканевой структуры и происхождением от внешне расположенных тканей — экзогенным происхождением (греч. ёхо— вне, снаружи и genesis — происхождение), тогда как настоящие корни закладываются эндогенно (греч. endon — внутри). У сфагновых мхов нет и ризоидов.
В ходе эволюционного развития корень и сте бель формировались одновременно и уже в заро дыше семенных растений зачаточный побег и корневой полюс составляют структурно целост ную ось растения. Их морфофизиологическое сходство свидетельствует о родстве. Следователь но, корневые системы нельзя анализировать в от рыве от системы побегов, причем и ту, и другую следует рассматривать в динамике. В учебной ли тературе органы растений рассматриваются как самостоятельные части организма для более пол ного и целостного представления об их структуре и функциях.
МОРФОЛОГИЧЕСКОЕ СТРОЕНИЕ КОРНЯ. КОРНЕВЫЕ СИСТЕМЫ
Корень закладывается уже в зародыше семени. Развивающийся из зародышевого корешка корень называют первичным. У голосеменных и двудоль-
163
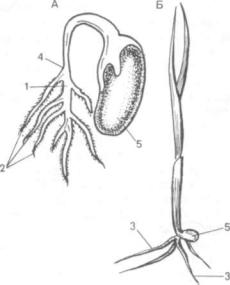
ных (особенно древесных — береза, дуб, груша и др.), многих травянистых (бобы, фасоль, горох, свекла) он становится главным корнем и растет прямо вниз. По мере роста главный корень зна чительно утолщается и разветвляется, образуя бо ковые корни, самые старые из которых находятся ближе к стеблю, а самые молодые ближе к кон чику корня (рис. 127, А, см. рис. 126).
Рис. 127. Развитие корня:
Л — у фасоли (Phascolus vulgaris); Б — у пшеницы (Triticum durum); l — глав ный корень; 2 — боковые корни; 3 — придаточные корни; 4 — корневая шейка; 5 — семя фасоли, зерновка пшеницы
Убольшинства двудольных растений главный корень бывает тоньше, чем стебель, поэтому пе реход стебля в корень выражен довольно четко. Этот участок перехода стебля в корень носит специальное название — корневая шейка (см. рис. 126, 127).
Уоднодольных первичный корень, как прави ло, недолговечен и формирующиеся корни берут начало от нижней части стебля (рис. 127, Б). Корни, берущие начало от других органов расте ния (стеблей, листьев, клубней, луковиц), на зываются придаточными или адвентивными (рис. 127, Б, 3). По мнению одних авторов, они нико гда не образуются ни от зародышевого корешка,
ни от |
главного корня (В. Б. |
Хржановский, |
1982; |
В. X. Тутаюк, 1980 и др.). |
Н. С. Воронин |
(1988) считает, что придаточные корни могут об разовываться и на старых участках корня (корнеродные придаточные корни). Ткани, в ко торых возникают придаточные корни, очень раз нообразны — апикальная меристема, камбий, феллоген, сердцевинные лучи и т. д. От прида точных корней, так же как от главного, отходят соответственно боковые корни — оси второго и последующих порядков ветвления. Таким обра зом, у однодольных развиваются в основном при даточные корни. При вегетативном размножении двудольных черенками, усами, отводками, клуб нями, корневищами развиваются только прида точные корни. У картофеля, например, при раз множении клубнями образуются придаточные
корни, а при размножении семенами формирует ся хорошо выраженный зародышевый главный корень.
Внешне придаточные корни не отличаются от обычных боковых корней и выполняют те же функции.
Развитие корней в онтогенезе приводит к формированию корневой системы. Под корневой системой понимают совокупность всех корней одного растения независимо от их происхожде ния и структуры. Корни и корневые системы очень разнообразны по происхождению, морфо логии, типам ветвления, расположению. Отсюда разные подходы к классификации корней. Чаще всего в основе их классификации лежат проис хождение (генетический принцип), форма и ха рактер ветвления (морфологический) и отноше ние к субстрату (экологический). Типы корней по происхождению уже назывались — главный, боковые и придаточные корни. Поскольку при любой классификации речь идет об особенно стях развития этих трех типов корней, они счи таются общеморфологическими признаками кор ня. Чаще всего выделяют три типа корневых сис тем.
1. Система главного корня — состоит из глав ного корня, его боковых корней первого, второго и последующих порядков. Н. С. Воронин (1988) относит сюда и придаточные корни, образующие ся на перечисленных корнях. По отношению к главному корню боковые корни расположены более или менее перпендикулярно. Главный ко рень (ось первого порядка) всегда выделяется центральным местоположением, большими диа метром и длиной, хотя боковые корни второго и даже третьего порядка по толщине мало уступают главному корню (рис. 128, А). Такая корневая система характерна для большинства голосемен ных и древесных двудольных, не образующих придаточных корней на стеблях, и многих трав, особенно однолетних. В некоторых случаях глав ный корень не разветвляется или ветвится ма ло — простой корень. Такой корень имеют кермек широколистный, морковь, ряска, а также ви ды с втягивающими корнями (лилии). Систему главного корня часто называют стержневой.
2. Система придаточных корней (рис. 128, Б, В). Придаточные корни развиваются в основном на стебле (стеблеродные придаточные корни).
У наиболее древних наземных растений (плаунов, хвощей, папоротников) главный корень вообще отсутствует и корневая система изначаль но формируется только из придаточных корней. Это явление, получившее название первичной гоморизии (греч. homos — одинаковый и rhiza — корень), наиболее примитивное в эволюционном отношении. Появление в процессе эволюции се мени, содержащего зародыш с зародышевым ко решком, привело к образованию системы главно го корня — аллоризии (греч. alios — иной, другой).
У однодольных главный корень рано редуци руется и формируется система придаточных кор ней. Такое явление получило название вторичной гоморизии (злаки — ячмень, овес, пшеница и др., осоки, лилейные). У двудольных система прида-
164
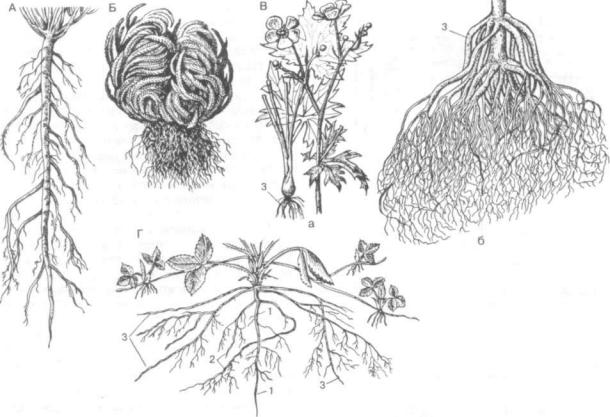
Рис. 128. Корневые системы:
Л — главного корня (аллоризная) одуванчика (Taraxacum officinale); Б — первично гоморизная у селагинеллы тамарисколистной (Sellaginella tamariscina); В — вторично гоморизная: а — у лютика едкого (Ranunculus асег); б — у пшеницы (Triticum); Г — главного и придаточных корней (смешанная) у земляники (Fragaria vesca): I — главный корень; 2 — боковые; 3 — придаточные корни
точных корней развивается редко, например у лютиков.
3. Систему главного корня с возникающими затем на стебле придаточными корнями часто на зывают смешанной. Она встречается у многих дву дольных травянистых растений (подсолнечник, фасоль, земляника и др.).
ФОРМЫ КОРНЕЙ. ЭКОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РОСТА КОРНЯ
Впределах трех типов корневых систем мор фологическое разнообразие корней по общей конфигурации, консистенции, способам ветвле ния и распространения в субстрате чрезвычайно велико.
Всистеме главного корня в зависимости от морфоструктуры основной оси бывают такие формы: цилиндрическая (тмин, хрен, кермек ши роколистный), веретенообразная (морковь), реповидная (редис, репа, редька), клубневидная (георгина, чистяк), древовидная (у многих дре весных, особенно в молодом состоянии).
Урастений с высоко расположенными под земными побегами (однолетние и плотнокустовые злаки, многие эпифиты) система придаточ ных корней по форме мочковатая, формирую щаяся у основания стебля.
По консистенции корни бывают мясистые, травянистые, одревесневающие.
По способам ветвления и внешнему виду вы деляют простые, разветвленные и нитевидные корни (мочковатые).
В зависимости от распределения массы кор ней по горизонтам почвы корневые системы бы вают поверхностными, глубинными и универ сальными, что обусловлено приспособлением растений к существованию на различных почвах и в условиях различного водоснабжения.
Пластичность растений, имеющая большое значение в процессе приспособления к различ ным почвенным условиям, во многом определя ется особенностями корневой системы.
При оптимальных условиях обитания корни растений проникают на большую глубину в почву и сильно разрастаются в ширину. Нередко расте ния образуют корневую систему в 2—3 яруса, что обусловлено расположением в почве питательных веществ и влаги (рис. 129, А). У хлебных злаков (пшеница, рожь, ячмень, кукуруза), корнеплодов корни могут проникать на глубину 1,5—2 м. Из травянистых растений длинную корневую си стему имеют люцерна (5—10 м и более), клевер (3 м). Но основная масса корней у однолетних растений сосредоточена в пахотном слое, на глу бине 30—40 см. У древесных растений корни проникают на глубину до 10—15 м. Рекордная глубина проникновения корней в почву отмечена у пустьшного мескитового кустарника (Prosopis juliflora): их находили на глубине 53,3 м в штате Аризона (США). Корни тамариска и акации об наруживались на глубине 30 м (Египет). Пло-
165
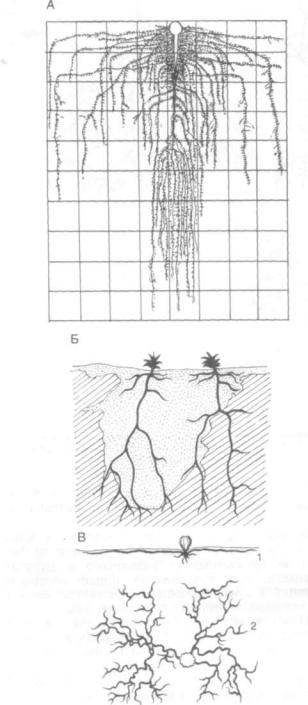
Рис. 129. Расположение корневой системы в почве:
А — свекла (Beta vulgaris); В — верблюжья колючка (Alhagi); В — кактус (Ferocactus vrislizenii): I — вид сбоку; 2 — сверху
щадь, занятая корневой системой, у древесных растений намного превышает диаметр кроны и достигает 10—18 м.
Обильное ветвление корней обеспечивает ог ромную всасывающую поверхность. Так, общая длина корневой системы у одного растения хлеб ных злаков, вытянутая в длину, включая корне вые волоски, будет равна 20 км. При специаль ной агротехнике длина всех корней риса дости гает 630 км; если к этой цифре добавить длину корневых волосков, то получится 11 000 км. При этом суточный прирост корней равняется при
мерно 5 км, а общая поверхность корней превы шает площадь надземной массы в 130 раз и более.
Темпы развития корней большинства расте ний превышают темпы развития надземных побе гов. Например, у проростка люцерны в фазе 2-3- го листа корень, разветвляясь, углубляется до 1 м. На той же фазе корень овса углубляется до 80 см, пшеницы — на 45, кукурузы — на 30 см. У дву летнего сеянца дуба высотой 30—40 см главный корень успевает углубиться на 1,5—2 м.
Корневая система растений обладает не толь ко большой длиной, она велика и по массе. Счи тается, что масса корней озимой ржи составляет 3—4 т/га, а многолетних травосмесей в пахотном слое — более 4—7 т/га.
Развитие корней связано с внешней средой. Особенно существенное влияние оказывает влага. У растений пустынь и полупустынь корни не только значительно углубляются, но могут обра зовывать два-три яруса. У саксаула в Каракумах корень образует мощно ветвящийся второй ярус на глубине 6—7 м. На тех же песках джузгун об разует корни другого типа — горизонтальные, за крепляющие подвижный субстрат: главный ко рень углубляется всего на 1,5—2 м, а горизон тальные — захватывают более 20 м. Мощная корневая система образуется и у травянистых рас тений пустынь и полупустынь. У верблюжьей ко лючки высота надземной части составляет 50— 60 см, а корни углубляются на 15—20 м к под земной влаге (рис. 129, Б). Корневая система многих кактусов широко разветвляется близ по верхности почвы (рис. 129, В). На болотных поч вах при избытке влаги корни растений развива ются главным образом в верхних слоях почвы, где они обеспечены достаточным количеством воздуха и влаги.
Почва, на которой произрастают растения, имеет первостепенное значение в развитии кор ней. На песчаных, легких почвах корневая систе ма проникает глубже в направлении грунтовых вод и хорошо ветвится. На глинистых почвах ко рень растет в глубину с большим трудом и в основном развивает боковые горизонтальные корни. В зоне вечной мерзлоты Крайнего Севера корневая система также развивается в горизон тальном направлении. Мощная корневая система развивается на черноземах, богатых питательны ми веществами. Некоторые растения, например можжевельник, инжир, в отдельных случаях бук, сосна, свободно растут и развиваются на скалах. Такие растения называются скальными. Особен ности корневой системы и высокое тургорное давление в процессе роста приводят к тому, что скалы раскалываются. Кислотные выделения корней постепенно растворяют субстрат.
В природных условиях корни травянистых растений различных видов располагаются поярусно и плотно переплетают всю толщу почвы.
Некоторые растения, предпочитая определен ный тип почвы, тем не менее способны расти на других субстратах. Так, сосны — растения песча ных почв — встречаются на болотах, скалах, в условиях тундры, в лесостепи и т. д. Такие расте ния, обладающие необычайной пластичностью
166
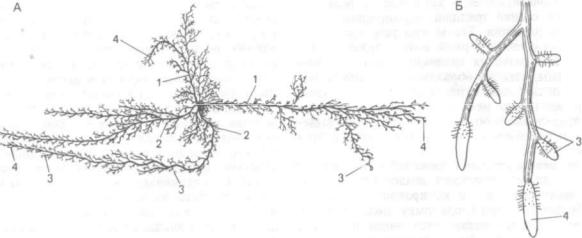
Рис. 130. Дифференциация корней в корневой системе яблони — Malus domestica (схемах):
А — общий вид корневой системы; Б — корневая мочка; 1 — скелетные; 2 — полускелетные корни; 3 — ростовые корни; 4 — сосущие окончания
корневой системы, часто называют растениямикосмополитами.
У культурных растений развитие корневой системы зависит от качества обработки почвы, удобрения, полива и ухода за растениями.
В зависимости от того субстрата, в котором расположены корни и откуда они извлекают воду и питательные вещества, различают четыре эко логических типа корней: подземные, водяные (или плавающие), воздушные и гаустории.
Подземные корни целиком или хотя бы час тично расположены в грунте. Такие корни имеют до 70 % высших растений. Они чрезвычайно раз нообразны по структуре и функциям.
Водяные корни всегда расположены в толще воды и никогда не достигают дна водоема (ряска, водокрас обыкновенный).
Воздушные корни находятся в воздухе, погло щая атмосферную влагу. Такие корни бывают у растений-эпифитов — обитателей влажных тро пических лесов.
Гаустории — корни-присоски паразитических растений (повилика, омела).
Перечисленные морфологические особенно сти дают некоторое представление о разнообра зии корневых систем. В любой корневой системе постоянно идут процессы отмирания, наращива ния, дифференциации, приводящие ее в соответ ствие с возрастом надземной системы побегов, сменой сезонов года, окружающими условиями. Без знания этих процессов нельзя понять, как взаимодействуют между собой растения леса, лу га, болота и других фитоценозов.
Типичная, нормально развитая корневая сис тема включает участки разной структуры и функ ций. У древесных растений различают толстые
скелетные и полускелетные корни, на которых возникают недолговечные обрастающие корневые мочки, развивающиеся близко к поверхности почвы, особенно в связи с дождями или поли вом. В состав корневых мочек, непрерывно за меняющих друг друга, входят ростовые и сосущие окончания. Ростовые окончания обычно более мощные по сравнению с сосущими, быстро уд линяются и продвигаются в глубь почвы. В них хорошо развита и энергично работает апикальная
меристема. Сосущие окончания, возникающие в большом количестве на ростовых корнях, растут медленно, и их апикальные меристемы часто почти прекращают свою деятельность. Сосущие окончания как бы останавливаются в почве и ин тенсивно поглощают питательные вещества дан ного участка (рис. 130). Сосущие корешки обыч но недолговечны. Ростовые корни могут длитель но существовать или отмирать вместе с сосущими корнями. Скелетные и полускелетные ветви часто запасают питательные вещества. У травяни стых растений в корневой системе в основ ном дифференцируются ростовые и сосущие корни.
Структурные изменения в корневых системах растений, используемых человеком, представляют интерес для растениеводов, поскольку почти все агротехнические приемы (вспашка, удобрение, полив, пересадка, прополка и др.) направлены в основном на создание оптимальных условий для развития и деятельности корневых мочек. В связи с этим ученые многих направлений (ботаники, луговеды, агрономы, лесоводы, физиологи, био химики) серьезно изучают корневую систему рас тений, разрабатывая разнообразные методы ее эффективной деятельности.
МЕТОДЫ ИЗУЧЕНИЯ КОРНЕВЫХ СИСТЕМ
Изучение корневых систем связано с больши ми трудностями. Динамику роста и развития кор ня можно изучить только на «работающей» кор невой системе, в естественных условиях ее оби тания. Выделение корней из почвы даже у тра вянистых растений связано с частичным их трав мированием, корни древесных растений полно стью извлечь из почвы вообще невозможно. К тому же в вынутых из почвы корнях быстро пре кращаются многие процессы жизнедеятельности, и они погибают.
Наиболее полное представление можно соста вить, сравнив данные, полученные разными ме тодами. Чаще всего для изучения корневых сис тем используют следующие методы.
167
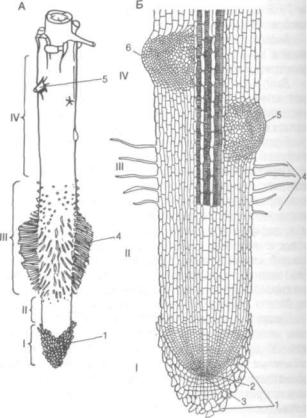
и выполняет ответственную функцию всасывания почвенных растворов. Этому способствует спе цифическое строение молодого корешка, неоди наковое на всем протяжении. Выделяют ряд уча стков, или зон, которые отличаются морфоанатомическими особенностями и выполняют различ ные физиологические функции (рис. 131).
Зоны корня. Кончик корня покрыт в виде на перстка корневым чехликом — особой паренхимной тканью, которая защищает апикальную ме ристему от трения о почвенные частицы и спо
Выращивание опытных растений в специальныхсобствует продвижению корня. В процессе роста
ящиках — ящики заполняют землей нужного со |
корня и проталкивания корневого чехлика вперед |
става, закапывают в землю, проводят специаль |
клетки чехлика на периферии слущиваются, об |
ные обработки почвы (подкормку, полив и т. д.). |
разуя слизь, которая обволакивает корень и об |
Корневая система развивается нормально при |
легчает его скольжение между частицами почвы. |
заданных условиях. Через определенное время |
Апикальная меристема образует все новые и но |
ящик выкапывают, фиксируют корневую систему |
вые клетки корневого чехлика, которые быстро |
длинными гвоздями, вколачивая их через боко |
отмирают (продолжительность их жизни 4—9 |
вую стенку, и отмывают струей воды, сняв про |
дней в зависимости от длины чехлика и вида рас |
тивоположную боковую стенку. Таким образом |
тения). Кроме того, они слабо соединены между |
можно изучить общую массу корней, распределе |
собой, что облегчает их слущивание. Углублению |
ние и соотношение их частей. |
корня способствует тургорное состояние клеток |
Сезонные и возрастные изменения в корневых |
чехлика. Центральная осевая часть чехлика со |
системах изучают, откапывая растения в разные |
ставляет так называемую колумеллу, в клетках ко |
сроки. |
торой содержится много крахмальных зерен. |
Наблюдение за ростом корней можно вести из |
Слизь, секретируемая наружными клетками |
специальных подвалов через стеклянные окна, |
чехлика, представляет собой сильно гидратиро- |
обращенные в почву. Однако вся корневая систе |
ванный полисахарид, вероятно, пектиновой при |
ма при этом не видна, а лишь те ее ответвления, |
роды. Она накапливается в пузырьках диктиосом, |
которые прижимаются к стеклу. |
|
В последние годы корневые системы изучают |
|
на растениях, выращиваемых в специальных |
|
стеклянных сосудах на питательных растворах. |
|
Корневая система при этом затеняется специаль |
|
ными плотными экранами. Так можно моделиро |
|
вать различный режим питания, наблюдать дина |
|
мику роста корней. |
|
В связи с техническими трудностями, возни |
|
кающими при исследовании корней, их строение |
|
и развитие изучены пока недостаточно. Тем не |
|
менее к настоящему времени в этой области име |
|
ется обширный научный материал по многим |
|
культурным и дикорастущим растениям, который |
|
используется в различных отраслях народного |
|
хозяйства (растениеводстве, плодоводстве, лесо |
|
ведении и т. д.). |
|
Происхождение и рост первичных тканей. Строение молодого корешка. Рост большинства корней представляет собой, по-видимому, непре рывный процесс, который затормаживается или прекращается только при неблагоприятных усло виях, таких как низкая температура и засуха. В почве корни распространяются по пути наи меньшего сопротивления и часто заполняют про странства, оставшиеся после ранее отмерших и сгнивших корней. Нередко корням приходится прокладывать путь в очень твердой почве. Не смотря на тесный контакт с твердыми почвенны ми частицами, растущий корень не повреждается
Рис. 131. Общий вид (А) и продольный разрез (схема, Б) моло дого корешка:
/— корневой чехлик, прикрывающий зону деления; II — зона растяжения; III — зона всасывания, или зона корневых волосков; IV — начало зоны проведения, или зоны боковых корней; 1 — корневой чехлик; 2 — группа инициальных клеток; 3 — калиптроген; 4 — корневые волоски; 5 — выход бокового корня; б — боковые корни
168
которые, сливаясь с плазматической мембраной, высвобождают слизь в пространство между плазмалеммой и клеточной оболочкой. Слизь затем выходит на поверхность оболочки, образуя мел кие капли.
Корневой чехлик есть на кончиках корней всех наземных растений, а у водных вместо чехлика формируется чехликоподобное образование — корневой кармашек. Если водное растение попадает в почвенные условия, на кончике его корней по является настоящий корневой чехлик. У многих однодольных в кончике корня хорошо развита меристема корневого чехлика, называемая калиптрогеном (чехликообразователь). По данным ис следователей, в калиптрогене синтезируются гор моны — ауксины, стимулирующие рост корня.
Корневой чехлик выполняет и другие важные функции, контролируя, в частности, реакцию корня на гравитацию (геотропизм).
Корневой апекс, или зона деления, находится внутри чехлика. Это апикальная меристема кор ня, его точка роста. Длина ее колеблется от 1 до 5 мм. Состоит из относительно мелких (10— 20 мкм в диаметре) многогранных клеток — инициалей и их непосредственных производных, отли чающихся густой цитоплазмой и крупными ядрами. На живом корешке эта зона хорошо заметна по бо лее темной (желтовато-бурой) окраске.
Хотя зона инициалей апикальной меристемы корня считалась когда-то местом активных кле точных делений, сейчас доказано, что это не так и большинство делений происходит на некотором расстоянии от инициалей. Инициальные клетки отличаются от остальных клеток меристемы более растянутым периодом деления, более медленным синтезом ДНК, РНК и белков, меньшим числом плазмодесм. Эти клетки получили название по коящегося центра. Область центра относительно неактивна, но при определенных условиях деле ния в ней происходят и покоящийся центр спо собен восстанавливать периферические зоны ме ристемы при их повреждении. Например, изоли рованные покоящиеся центры кукурузы, выра щенные в стерильной культуре, способны вырас тать в целые корни, минуя стадию каллуса, или раневой ткани; обнаружена четкая корреляция между размерами покоящегося центра и сложно стью строения первичной проводящей системы корня кукурузы. Результаты исследований свиде тельствуют о существенной роли покоящегося центра в развитии и формировании корня.
Зона растяжения (роста) находится дальше от апекса. Прекратившие деление клетки переходят к растяжению в направлении к оси корня. Имен но рост клеток в этой зоне обусловливает основ ное удлинение органа. Объем корня увеличивает ся за счет общего оводнения клеток и появления крупных вакуолей. Протяженность зоны роста не превышает нескольких миллиметров, внешне ко рень выглядит гладким и более прозрачным.
Зона всасывания следует за зоной роста. Клет ки резко прекращают растяжение, начинается их дифференциация. На покровной ткани (ризодермис) возникают многочисленные выросты — кор невые волоски, плотно охватывающие частицы
почвы и как бы «срастающиеся» с ними. Эту зону часто называют зоной корневых волосков или зоной дифференциации (созревания), поскольку наряду с выростами ризодермиса здесь начинают формироваться первичные ткани корня и осуще ствляться его основная функция — всасывание почвенных растворов. Опыты показали, что по давляющую массу воды и солей молодые корни воспринимают именно в этой зоне, имеющей протяжение до нескольких сантиметров.
Зона всасывания непрерывно передвигается в почве, так как старые корневые волоски отмира ют, а новые формируются со стороны зоны роста. Нарастание корня сопровождается также включе нием старых участков зоны всасывания в выше расположенную зону. Таким образом, всасываю щий аппарат молодого корешка — подвижное образование, непрерывно меняющее место в поч ве.
Зона проведения обладает хорошо развитой проводящей тканью и передает почвенные рас творы выше по органу. Корневые волоски вместе с клетками ризодермиса погибают, и формирует ся покровная ткань, защищающая живые ткани корня. В этой зоне появляются боковые корни, поэтому ее часто называют зоной боковых кор ней.
Важно отметить, что переход от одной зоны к другой происходит постепенно, без резких гра ниц. Некоторые клетки начинают удлиняться и дифференцироваться еще в зоне клеточных деле ний, в то время как другие достигают зрелости в зоне растяжения. Так же закономерно, в опреде ленной последовательности, возникают в моло дом корешке внутренние ткани.
Апикальный рост корней. Корень отличается от побега тем, что его апикальная меристема откла дывает клетки внутрь и наружу, формируя чех лик. Производные апекса, откладываемые внутрь корешка, образуют хорошо заметные в структуре кончика корня продольные ряды клеток.
Организация апикальной меристемы (число инициалей, порядок их деления) у разных систе матических групп растений неодинакова. У мно гих папоротниковидных апекс корня состоит из одной инициальной, или верхушечной, клетки тетраэдрической формы, дающей начало всем клеткам и тканям корня (рис. 132, А). Деление инициальной клетки происходит параллельно каждой грани тетраэдра. После каждого последо вательного деления инициальная клетка восста навливает свои форму и размеры. Отложенные ею клетки, многократно делясь, образуют четко оформленные комплексы.
В корнях семенных растений наблюдается не сколько типов апикальной организации. У по крытосеменных апекс корня многоклеточный. При этом строение и функционирование апи кальной меристемы у двудольных и однодольных имеют существенные различия.
У двудольных растений в типичном случае инициальные клетки расположены тремя слоями, каждый из которых содержит 1—4 инициали (рис. 132, Б, б). Инициали нижнего слоя дают начало чехлику и ризодермису. Инициали сред-
169
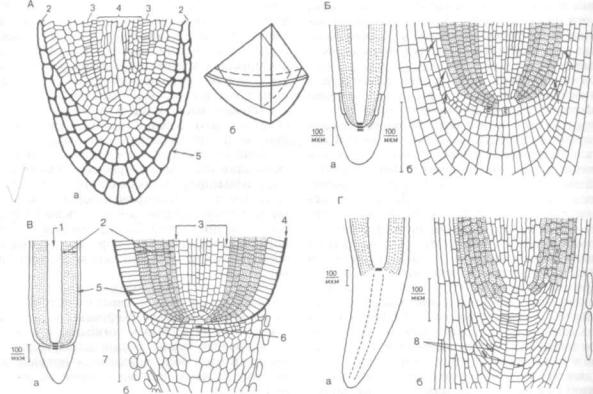
Рис. 132. Апикальная организация корня в продольных разрезах (по К. Эсау, 1969, 1980):
А — папоротника деннштедтия веховидная (Dennstaedtia cicutaria): a — организация апекса с верхушечной клеткой; б — объемная схема деления верхушечной клетки; / — верхушечная клетка; 2 — ризодермис; 3 — эндодерма; 4 — центральный цилиндр; S — корневой чехлик; Б, а, б — ре диса (Raphanus). Видны три зоны организации. Ризодермис и корневой чехлик имеют общее происхождение; В, а, б — злака ковыля (Stipa). Видны три зоны инициалей, среди них инициали чехлика, формирующие калиптроген. Эпидермис и кора имеют общее происхождение; Г, а, б — ели (Ксеа). Все зоны корня происходят из одной группы инициалей. Корневой чехлик имеет центральную колонку, 1 — центральный ци линдр; 2 — первичная кора; 3 — перипикл; 4 — слизистый слой; 5 — эпидермис; б — калиптроген; 7 — корневой чехлик; 8 — колонка
него и верхнего слоев образуют все остальные ткани собственно корня. На рис. 132 видно, что уже в зоне деления, т. е. в самой апикальной ме ристеме, обнаруживается четкое разграничение на три зоны: наружная зона — дерматоген (греч. derma, род. п. dermatos — кожа и genes — рожден ный). Клетки дерматогена относительно правиль ной формы, делятся антиклинально, образуют ризодермис с корневыми волосками. У двудоль ных вместе с калиптрогеном участвуют также в формировании корневого чехлика. Средний отдел инициалей формирует периблему (греч. periblema — оболочка, покров), из которой впоследст вии возникают клетки первичной коры. Деление клеток в периблеме идет во всех направлениях, форма их неправильная. Внутренняя зона, кото рая берет начало от верхнего слоя инициалей, называется плеромой (греч. pleroma — наполне ние). Ее клетки несколько вытянуты по оси, де лятся преимущественно антиклинально, из них формируется осевой, или центральный, цилиндр.
У однодольных типичный корневой апекс от личается от апекса двудольных тем, что нижние инициали образуют только чехлик, а ризодермис дифференцируется из самого наружного слоя пе риблемы (рис. 132, В, в).
Однако среди однодольных и среди двудоль ных растений встречаются виды, у которых апи кальная организация не дифференцирована на инициальные слои. Вся группа инициалей дает начало всем тканям корня, включая чехлик.
У голосеменных все зоны корня происходят из одной группы инициалей. Функцию дерматогена выполняет протодерма, которая дифференцирует ся из периблемы и формирует в дальнейшем ри зодермис и клетки корневого чехлика. Корневой чехлик имеет центральную колонку поперечно делящихся клеток. От этой колонки в латераль ном направлении возникают производные клетки (рис. 132, Г, г).
Представление о происхождении тканей корня из обособленных инициалей апекса было обосно вано в теории гистогенов Дж. Ганштейном (1868, 1870). Согласно этой теории, тело растения раз вивается из массива меристемы, которая состоит из многих слоев клеток и содержит три гистогена (этажа, уровня, слоя) — три предшественника тканевых зон. При этом каждый из гистогенов берет начало в апексе от одной или нескольких инициалей, расположенных ярусами друг над другом. Этими гистогенами являются дерматоген (предшественник эпидермиса), плерома (предше ственник центрального цилиндра) и периблема (предшественник первичной коры). Подразделе ние на три гистогена не имеет универсального применения, так как оно редко выявляется в по бегах, но для описания дифференциации ткане вых зон в корне удобно и часто используется.
Развитие постоянных тканей в корне. Диффе ренциация постоянных тканей в корне начинает ся очень близко к кончику корня. Вблизи апи кальной меристемы уже можно различить слои
170
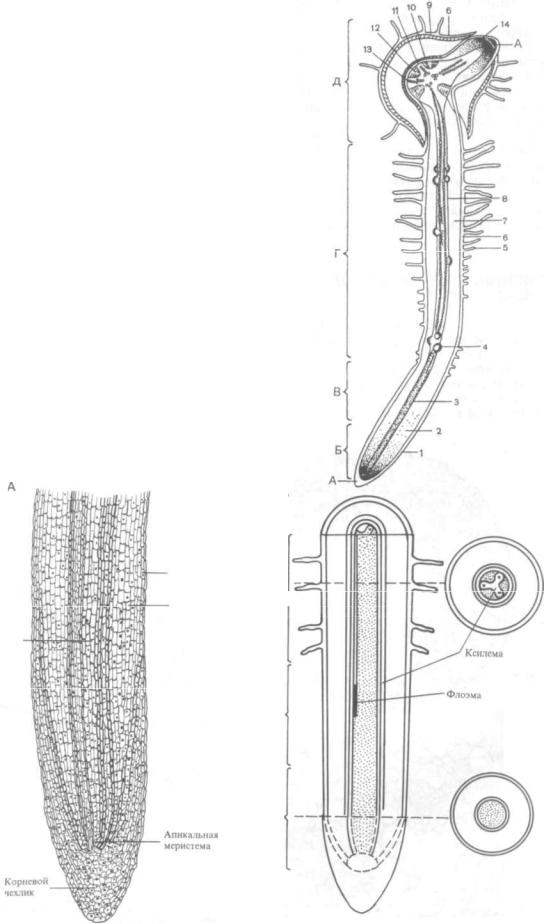
детерминированной меристемы — протодерму, прокамбий и основную меристему, т. е. первич ные меристемы, преобразующиеся соответственно в ризодермис, первичные проводящие ткани и первичную кору. Клетки — производные апи кальной меристемы — сначала удлиняются, затем дифференцируются в постоянные ткани в соот ветствии со своим положением в корне.
Первыми в центральном цилиндре обособля ются материнские клетки сосудов метаксилемы. Они занимают центральное положение. Но завер шается дифференциация в обратном порядке — первыми оформляются самые наружные сосуды, примыкающие к перициклу. Дальнейшее разви тие первичной ксилемы идет в центростреми тельном порядке. Элементы первичной флоэмы дифференцируются раньше, чем элементы ксиле мы. Это происходит еще в зоне растяжения кле ток. Флоэма также развивается центростреми тельно.
Ткани коры образуются из основной коровой меристемы. Наиболее активно делятся внутрен ние клетки коровой зоны, прилегающие к пери циклу. Новые клетки появляются в результате тангенциальных делений.
Рис. 133. Формирование первичных тканей в молодом корешке тыквы (Cucurbita pepo):
А — корневой чехлик; Б — зона деления; В — зона растяжения; Г — зона всасывания; Д — зона ветвления (проведения); 1 — дерматоген; 2 — периб лема; 3 — плерома; 4 — меристематические зачатки боковых корней; 5 — кор
невой волосок; |
6 — ризодермис; |
7 — кора; S — центральный цилиндр; |
Р — экзодерма; |
10 — эндодерма; |
11 — перипикл; 12 — первичная флоэма; |
13 — первичная ксилема; 14 — боковой корень
Протодерма
Зона Основная всасывания * меристема
Прокамбий
Зона
растяжения
Зона
Рис. 134. Формирование постоянных тканей в корне:
А — продольный срез кончика корня лука (АШшп сера); Б — схема стадий первичного роста кончика корня