

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfПаренхимные клетки воздухоносной ткани могут иметь различные модификации и сочета ния, что обусловливает характер межклетников. В одних случаях крупные паренхимные клетки звездчатой формы образуют перемычки и между ними остаются большие полости, заполненные воздухом (камыш озерный, ситник, рис. 115 А), в других — небольшого размера паренхимные клет ки, располагаясь цепочкой, окружают воздушную полость (рдест блестящий, кубышка, белокрыль ник, рис. 115, Б).
Аэренхима хорошо развита у растений, кото рые обитают в среде, затрудняющей нормальный газообмен и снабжение внутренних тканей ки слородом, например у водных и болотных расте ний. У многих из них она простирается от корня по стеблю до листьев. Помимо аэрации, воздуш ные полости внутри стебля и в листьях позволя ют растению свободно плавать в воде. Аэренхима выполняет также механическую (арматурную) функцию: ее структура, напоминающая пчелиные соты, наиболее полно и экономно обеспечивает прочность и эластичность органов тела растений в водной среде.
ВСАСЫВАЮЩИЕ ТКАНИ
Всасывающие ткани у растений довольно многообразны и играют важную роль в irx жизне деятельности, обеспечивая поступление воды и растворенных веществ. Они очень различны по структуре и по распространенности среди выс ших растений. Наиболее типично эта ткань пред ставлена в ассимилирующей зоне, молодого ко решка ризодермой (греч. rhiza — корень и derma — кожа) — наружным слоем клеток с выростами — корневыми волосками. Через ризодерму в корень из почвы всасывается вода с растворенными ве ществами (корневое питание).
Остальные типы всасывающих тканей, как правило, связаны с определенными условиями существования или приурочены к какому-то так сону. Поэтому более подробно они будут рас смотрены при изучении соответствующих орга нов или групп растений. Всасывающая ткань присосок (гаусторий) хорошо развита у растенийпаразитов, а также у насекомоядных и сапрофитов, веламен — на воздушных корнях орхидей. Поглощающие ткани развиваются в прорастаю щих семенах (например, на щитке у зародышей злаков) и в водопоглощающих волосках листьев.
У некоторых водных растений известны гидроподы, состоящие из одной или группы клеток, которые способны избирательно поглощать рас творенные в воде вещества (например, кубышка желтая).
К поглощающим тканям можно отнести ри зоиды многих низших растений, а также мико ризу.
Таким образом, в теле растительного организ ма, во всех его органах размещаются паренхим ные ткани, различающиеся физиологически, по этому даже в пределах одного органа могут быть функционально разные паренхимы. Однако все
паренхимы топографически могут быть класси фицированы по органам: паренхима корня (первичная и вторичная), паренхима стебля (первичная и вторичная), паренхима листа, пло да, семени и т. д.
ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
В отличие от животных, обмен у которых все гда связан с непрерывным выделением азотистых продуктов распада, у растений происходит реути лизация, или накопление, как поступающих, так и синтезируемых веществ в течение всей жизни с частичными потерями в виде листопада, веткопада, слущивания наружных слоев корки и т. п. Многие древесные растения в течение сотен, а то и тысяч лет сохраняют органические вещества в виде отмерших тканей, не прерывая роста и наращивая фитомассу.
В то же время многие вещества, которые про дуцируются растительной клеткой и, повидимому, являются побочными продуктами ме таболизма, не используются растением, а изоли руются от живого протопласта или удаляются из тела растения. Эти вещества (секреты или экс креты) накапливаются или выбрасываются из тела растения особыми тканями — выделительны ми, которые также часто называют секреторными структурами.
Функция секреции свойственна в той или иной степени любой живой растительной клетке. У выделительных тканей, представленных оди ночными клетками (идиобласты) или клеточными комплексами, секреция является преобладающей функцией.
Выделительные ткани изучены пока недоско нально, классификация их довольно противоре чива. Связано это прежде всего с огромным раз нообразием их структуры и размещения в расти тельном организме. Секретируемые растением вещества имеют разную химическую природу, образуются растением в различные периоды онтогенеза; причем одни и те же выделительные ткани могут продуцировать различные секреты, а одинаковые вещества выделяются разными типа ми секреторных структур. Функция многих выде ляемых веществ в жизнедеятельности растений до сих пор не выяснена, можно только предполагать, что они образуются при наиболее интенсивных обменных процессах, сопровождающих рост, за кладку новых органов, размножение. Достоверно изучены функции лишь немногих выделительных структур — пищеварительных железок, нектарни ков.
Накопление у многих растений в значитель ных количествах катаболитов (например, смол, эфирных масел) можно объяснить определенны ми дополнительными функциями, которые эти вещества-отбросы получили в ходе адаптивной эволюции. Например, наличие смол в древесине хвойных придает ей большую устойчивость про тив гниения, защищает от поедания животными.
Вещества-секреты представлены несколькими группами. Прежде всего это терпены (сильно
152
восстановленные производные непредельного углеводорода изопрена). С увеличением конден сационного числа (п равно от 500 до 5000) обра зуются все менее летучие жидкости, затем твер дые вещества (эфирные масла — бальзамы — смолы — каучук). Терпены чаще всего находятся в смеси друг с другом. Например, живица (тер пентин) — смолистое вещество, которое выделя ется при ранении хвойных деревьев, содержит 40—65 % смоляных кислот, 20—35 % монотерпе нов, 5—20 % сесквитерпенов и дитерпенов. Жи вица — основное сырье для получения канифоли и скипидара, которые представляют собой смеси химических соединений.
Кроме того, растения секретируют белковые вещества, полисахариды (сахара, слизи), соли, воду. Характерные черты большинства секретор ных клеток в активном состоянии: мелкие разме ры, густая цитоплазма, наличие тонких неодревесневающих оболочек, отсутствие в большинст ве случаев хлоропластов, а поэтому гетеро трофный характер метаболизма.
Электронно-микроскопические исследования показали высокую насыщенность цитоплазмы секреторных клеток органоидами определенного типа, связанными с химической природой выде ляемого вещества. Так, выделение белков связано с сильным развитием гранулярного, а выделение терпенов — агранулярного эндоплазматического ретикулума, продуцирование слизи — с развити
ем комплекса Гольджи. В связи с этим описание ультраструктуры секреторных тканей проводится по основному выделяемому веществу.
Секреторные ткани весьма разнообразны по морфологии и топографии в теле растения. Секретируемые продукты у идиобластов накаплива ются в центральной вакуоли или в пространстве между плазмалеммой и оболочкой. Примерами могут служить кристаллоносные клетки, содер жащие кристаллы оксалата кальция, эфирномасляные клетки, слизевые идиобласты, синтези рующие и выделяющие аморфные полисахариды.
Классификация секреторньгх комплексов (довольно условная) основана на их микрострук туре и расположении. Одни из них располагаются на поверхности надземных частей растения и выделяют секрет наружу — наружные выдели тельные ткани (железистые волоски и многокле точные железки); другие локализованы внутри органов, не связаны непосредственно с наруж ной средой и имеют вид каналов (ходов) или полостей, выстланных эпителием — внутренние выделительные ткани.
НАРУЖНЫЕ ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
Наружные (поверхностные) секреторные струк туры характеризуются общим происхождением. Все они возникают либо из клеток эпидермиса,
Рис. 116. Железистые волоски:
А — чашелистика айвы обыкновенной (Cydonia oblonga); Б — плода аниса обыкновенного (Anisiura vulgare): I — волосок с бородавчатой кути кулой; 2 — эфирномасляные канальцы; В — многоклеточный железистый волосок (7) и эфирномасляная железка (2) листа багульника болот ного (Ledum palustre): 2а — внешний вид железки; 26 — в разрезе видны капли эфирного масла; Г — железки на листьях березы бородавчатой (Betula verrucosa): a — внешний вид, б — поперечный срез, в — бурые клетки с накопленным суберином; Д — железки на листьях яснотки бе лой (Lamium album): 1 — кутикула, отодвинутая секретом железки; Е — железистый волосок чашелистика земляники (Fragaria vesca) с однокле точной головкой; Ж— волоски семядолей какао (Theobroma cacao); 3 — масляная железка прицветника конопли посевной (Cannabis saliva); И — железка чашелистика костяники (Rubus saxatilis); К — железистый волосок смородины красной (Ribes rubrum); Л — пельтатная железка листа смородины черной (R. nigrum)
153
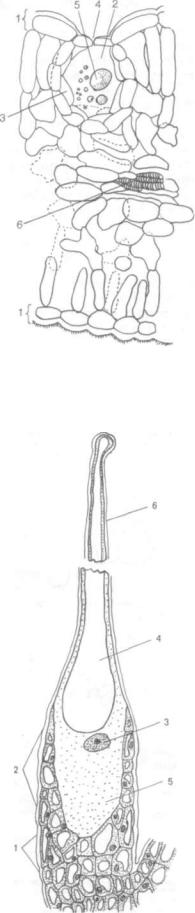
Рис. 117. Эфирномасляные железки листьев руты душистой (Ruta graveolens), поперечный срез листа:
) — эпидермис; 2 — железка; 3 — эпителиальные клетки железки; 4 — щель между «замыкающими клетками» железки; 5 — капли масла; 6 — проводящий пучок
Рис. 778. Жгучий волосок листа крапивы (Urtica dioica):
1 — основание волоска; 2 — жгучая клетка; 3 — ядро; 4 — вакуоль; 5 цитоплазма; 6 — обломившийся кончик жгучей клетки
154
либо из субэпидермальнъгх слоев. Продукты вы деления — эфирные масла, нектар, вода.
Железистые волоски. Они могут быть трихо мами и эмергенцами, содержащими пространство (вместилище) для накопления секрета и выдели тельную систему. Могут выводить из тела расте ния разнообразные секреторные вещества в газо образном, жидком, твердом виде, т. е. являются вьщелительными волосками. По строению мно гообразны, но видоспецифичны.
Более сложные секреторные структуры назы вают железками, однако резкой разницы между железками и железистыми волосками нет.
Железки можно классифицировать по место положению — наружные и внутренние; строе нию — однорядные, двухрядные, головчатые с од ноклеточной головкой и с многоклеточной го ловкой и ножкой; по продуктам выделения — сли зевые, масляные, камедевые, солевые (рис. 116).
Иногда наружные железки имеют вид щитка на ножке, например пельтатные железки у смо родины (рис. 116, Л). Железистые волоски обыч но выделяют секрет в пространство между обо лочкой и кутикулой; при этом кутикула сильно растягивается (рис. Мб, Д) и в конце концов ло пается. Иногда кутикула восстанавливается, на копление секрета происходит повторно. У неко торых растений от железистых волосков посте пенно отделяются верхние клетки с накопленным секретом, что, по-видимому, связано с гидрата цией и набуханием срединной пластинки.
Высокоспециализированные железки многих растений имеют довольно сложное строение. Так, у руты душистой железки листа погруженные (внутренние), представляют собой сферические схизо-лизигенные эфирномасляные вместилища, выстланные эпителиальными клетками. С внеш ней средой они сообщаются при помощи 2—4 «замыкающих» клеток, между которыми можно наблюдать очень узкую щель (рис. 117). Такое же строение железок у цитрусовых.
К наружным железистым эмергенцам можно отнести жгучие волоски. Они встречаются на над земных органах у некоторых представителей та ких семейств двудольных растений, как крапив ные (Urticaceae), молочайные (Euphorbiaceae), гидрофилловые (Hydrophyllaceae). Состоят из удлиненной, так называемой жгучей клетки, за остренной на конце, и многоклеточного основа ния. В вакуолях жгучей клетки синтезируются и накапливаются токсины (ацетилхолин, гистамин и др.). При соприкосновении, например, с шер стью животных вытянутый кончик волоска, имеющий пропитанные кремнеземом очень хруп кие стенки, обламывается и острыми краями впивается в кожу, выдавливая сок (рис. 118). Жгучий секрет раздражает кожу человека или животного, вызывая сокращение гладких мышц. Характерной особенностью жгучей клетки явля ется крупное (25—35 мкм), очевидно, полипло идное ядро, образующее глубокие лопасти, меж ду которыми встречаются пластиды и крупные вакуоли. В цитоплазме видны многочисленные митохондрии, сильно развит эндоплазматический ретикулум.
яг
Нередко наружные секреторные структуры представлены участками железистой ткани. Так, у смолки липкой (Viscaria viscosa, из сем. гвоздич ных) на стеблях под узлами образуются клейкие пояса. Эпидермис этих поясов дифференцирует выделительные клетки — крупные, богатые про топластом и снабженные сосочковидными высту пами наружной оболочки. В почечных чешуях многих древесных пород (тополь, береза и т. д.), некоторых трав (щавель) из клеток эпидермиса образуются крупные железистые участки, выде ляющие камеди, смолы, слизи и т. д.
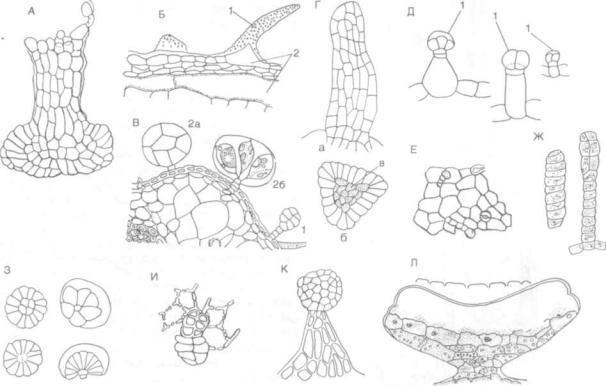
Солевые железки образуются на листьях, стеб лях многих растений солончаков — галофитов. Они служат для выведения (против градиента концентрации) на поверхность растения избытка минеральных солей, поступающих через ксилему из почвы, что позволяет растениям переносить очень сильное засоление субстратов. Так, у рас тений сем. плюмбаговых (Plumbaginaceae) — виды родов кермек, акантолимона и др. — солевые железки представляют многоклеточные образова ния, несколько напоминающие устьица, но более сложно устроенные (рис. 119). Это типичные эпидермальные трихомы, не связанные с прово дящей системой органов, на которых они распо ложены. На поверхности надземных частей их много (до 600—700 на 1 см2), секреция солей в них чрезвычайно интенсивная. По опытным дан ным, выделяемая за сутки листьями кермека ши роколистного жидкость составляет около полови ны массы самих листьев. Солевые железки сем. плюмбаговых получили даже особое название — «органы Ликбполи», по имени первого их иссле дователя, итальянского ботаника Гаэтано Ликополи. В последнее десятилетие появилось до вольно много работ о секреции солей и о солевых железках у галофитов (исследования голландских ботаников, работы А. Е. Васильева и др.).
Некоторые исследователи не выделяют соле вые железки в особую группу секреторных обра зований, а рассматривают их как водяные желез ки, или гидатоды, по сходству структуры и функ ции; концентрация солей в растворах бывает
различной, вплоть до очень малой, и тогда желез ки действуют в качестве типичных гидатод.
Гидатоды (греч. hydor, род. п. hydatos — вода
и hodos — путь). Это водяные устьица, комплекс клеток в листьях, обеспечивающий выделение из растений капельно-жидкой воды и солей. Этот процесс обычно называют гуттацией (лат. gutta — капля).
В гидатодах развивается группа рыхлых тонко стенных клеток мезофилла листа — эпитема. С эпитемой соприкасается ксилемная часть прово дящего пучка, в основном трахеиды (рис. 120, А). Водяные устьица эпидермиса, лежащего над гидатодой, отличаются тем, что щели их постоянно открыты. У многих растений это объясняется тем, что клетки водных устьиц рано теряют содержи мое. От хлоренхимы листа гидатода отделяется обкладкой, состоящей из крупных бесцветных клеток паренхимы. Иногда гидатода представляет собой многоклеточный волосок, например у фа соли — Phaseolus multiflorus; у некоторых расте ний они сложные и, по-видимому, связаны с секреторной тканью.
Гуттацию можно наблюдать у многих расте ний, особенно у травянистых (земляника, ман жетка, буквица), рано утром или в условиях по вышенной влажности. Так как гидатоды чаще всего расположены по краям листьев, на зубчи ках, то как раз на них можно наблюдать висящие капельки воды. Иногда водные устьица распола гаются напротив разветвленных жилок (тутовые, крапивные). У камнеломок гуттационная во-
25 мнм |
|
|
|
Рис. 119. Солевая железка листа кермека Гмелина (Limonium |
|
|
|
gmelinii) на поперечном срезе: |
Рис. 120. Гидатода листа первоцвета (Primula sinensis): |
||
1 — секреторная клетка; 2 — пора в кутикуле; 3 — побочная клетка; 4 — |
А — продольный разрез; |
Б — вид с |
поверхности; 1 — эпидермис |
собирательная клетка; 5 — клетка эпидермиса; 6 — бокальчатая клетка; 7 — |
листа; 2 — замыкающая |
клетка водяного |
устьица; 3 — эпитема; 4 — |
мезофилл; 8 — кутикула |
хлоренхима; 5 — межклетники; 6 — проводящий пучок; 7 — обкладка |
||
155
шшшш^шшшшш
да, испаряясь, оставляет на листе осаждаю щуюся в твердом виде углекислую известь. У некоторых растений (смородина, черемуха, на стурция, эконит) с возрастом листа гидатоды за сыхают.
Значение гуттации, вероятно, заключается в том, что растение освобождается от избытков воды и солей. Выделяемая вода содержит также сахара и другие органические вещества.
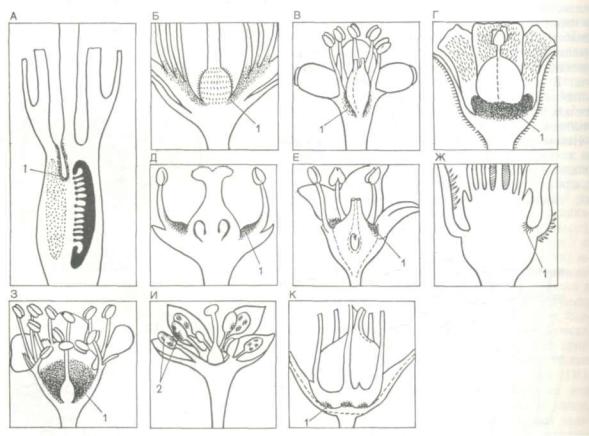
Нектарники (nectaria). Представляют собой разнообразные железистые образования, выде ляющие нектар. Располагаются большей частью в цветках (флоралъные нектарники), реже — на веге тативных органах (экстрафлоралъные, или внецветковые, нектарники). Флоралъные нектарники у двудольных могут находиться у основания ты чинок (гвоздичные, маревые); в виде диска или кольца у основания завязи (пасленовые, яснотковые); на вершине завязи в виде особой трубчатой структуры (астровые); на особых органах — медо виках. Иногда нектарниками являются видоизме ненные тычинки, или стаминодии (сем. барбари совых, мотыльковых). У однодольных нектарники часто находятся в выемках, или септах, завязи и имеют вид кармашков с железистой внутренней поверхностью. Если они глубоко погружены в завязь, то имеют выходные каналы, ведущие к поверхности завязи (рис. 121). Внецветковые нек
тарники могут развиваться на цветоножках, при листниках, стеблях и листьях.
Секреторная ткань нектарников формируется либо эпидермисом, либо субэпидермальной тка нью. К секреторной ткани обычно примыкает проводящая. Выводится нектар через особые устьица или непосредственно через стенку клетки при разрыве кутикулы.
В некоторых семействах (барбарисовые, люти ковые) отмечены определенные направления эво люции нектарников, например, от шпорцевых нектарников у видов аконита к лепестковидным у живокости, что ведет к увеличению околоцветни ка. В некоторых случаях сложное строение медо виков и нектарников с трудом поддается объяс нению, поскольку связь с насекомыми-опы лителями давно утратилась.
Нектар, продуцируемый этими секреторными структурами, — сахаристый сок, по составу — водный раствор Сахаров с небольшой примесью спиртов, азотистых и ароматических веществ, кислот, ферментов. Количество нектара, выде ляемое отдельным цветком за время цветения, у разных растений различно. У известного медо носного растения липы — от 0,15 до 7,46 мг, у малины — в среднем 14 мг. Функция нектара - привлечение насекомых и птиц, осуществляющих перекрестное опыление. Кроме того, нектар со-
Рис. 121. Флоралъные нектарники на продольных разрезах цветков (по К. Эсау, 1969):
А — нарцисса (Narcissus) в септе завязи; Б — наружный чая (Thea) в основании тычинок; В — кокколобы ягодоносной (Coccoloba uvifera) в виде колец под тычинками; / — сем. молочайных (Euphorbiaceae) в виде дисков под завязью; Д — сем. бересклетовых (Celastraceae) в виде дисков между завязью и тычинками; Е — сем. зонтичных (Umbelliferae) в виде дисков в верхней части ниж ней завязи; Ж— джута (Corchorus) в форме подушковидных собраний волосков; 3 — сливы (Pranus), выстилающий цветочную ча шу, И — коричника (Cinnamomum) в виде измененных бесплодных тычинок — стаминодиев; К — льна (Llnum) в виде железок у основания тычинок (1 — нектарники; 2 — стаминодии)
156
„*са,.^

держит вещества гормональной природы, участ вующие в репродуктивных процессах (прораста ние пыльцы, оплодотворение, развитие завязи). Нектар обладает также бактериостатическими свойствами. Из собранного с цветков нектара пчелы вырабатывают мед.
У волчеягодника, багульника, азалии, чемери цы нектар ядовитый.
Осмофоры (греч. osme — запах и рпоге — но ситель). Присущий многим цветкам аромат, при влекающий насекомых-опылителей, обусловлен летучими веществами, главным образом эфирны ми маслами, которые находятся в клетках эпи дермиса лепестков. Однако у некоторых долгоцветущих растений (сем. орхидные, ластовневые, аристолоховые) аромат продуцируют особые же лезки, названные С. Фогелем (1949) осмофорами. Различные части цветка могут дифференциро ваться в осмофоры; при этом они могут быть крыловидные, иногда в виде ворса, ресничек (рис. 122). Секреторная ткань осмофор обычно многослойная, выделение летучего секрета про исходит в течение короткого времени и связано с использованием большого количества запасных веществ.
Кроме специализированных образований ос мофор, запахи источают лепестки, тычинки, ста минодии, нектарники, пыльца. Причем разные части цветка имеют специфические запахи и на секомые хорошо их различают.
Летучие эфирные масла, которые в основном придают характерный аромат цветкам и их частям у различных видов растений, по физическим и химическим показателям весьма разнообразны. Компонентами эфирных масел являются спирты, альдегиды, фенолы и другие вещества. Так, аро мат цветка розы обусловлен первичными спирта ми (гераниолом, перолом, цитронеллом), а запах
Рис. 122. Осмофоры в цветках растений (по К. Эсау, 1969):
а — спартиум (Spartium junccum); б — любка двулистная (Platanthera bifolia); в — нарцисс (Narcissus jonguilla); г — люпин (Lupinus crackshanksii); д - дендробиум (Dcndrobium minak). Осмофоры показаны точками
фиалки — кетонами, пармонами. Различают око ло 500 разных цветочных запахов. Эфирные масла придают запах вегетативным органам, чаще ли стьям, многих ароматических, пряных растений. Распространяются они, как и газы, токами воздуха.
К наружным выделительным тканям относят
пищеварительные железки насекомоядных расте ний (росянки, пузырчатки и др.). Жидкость, вы деляемая этими железками, содержит вещества (ферменты, кислоты), с помощью которых пере вариваются пойманные животные.
ВНУТРЕННИЕ ВЫДЕЛИТЕЛЬНЫЕ ТКАНИ
Внутренние выделительные ткани представле ны у растений идиобластами, особыми вмести лищами выделений и млечниками. Секрет, нака пливаемый этими тканями, может храниться на протяжении многих лет и выводится во внешнюю среду только при механических повреждениях или при естественном отмирании и опадении органов (листьев, веток и т. д.). Продуктами внутренней секреции служат смолы, эфирные масла, дубильные вещества, кристаллы мине ральных солей и др.
Выделительные клетки — идиобласты. Встре чаются среди клеток разных тканей, могут иметь разнообразную форму и химический состав. Они накапливают различные вещества — кристаллы оксалата кальция (одиночные, друзы, рафиды и т. д.; см. рис. 59), слизи, таннины, эфирные мас ла. Последние начинают накапливаться в выде лительной клетке внутри пузырька, прикреплен ного к оболочке. Постепенно пузырек разрастает ся и замещает полость клетки. Одновременно на оболочку клетки изнутри откладывается суберин, который изолирует ядовитое содержимое от ок ружающих тканей.
Эфирномасляные идиобласты характерны для ряда семейств — магнолиевых, перечных, лавро вых, кирказоновых и др.
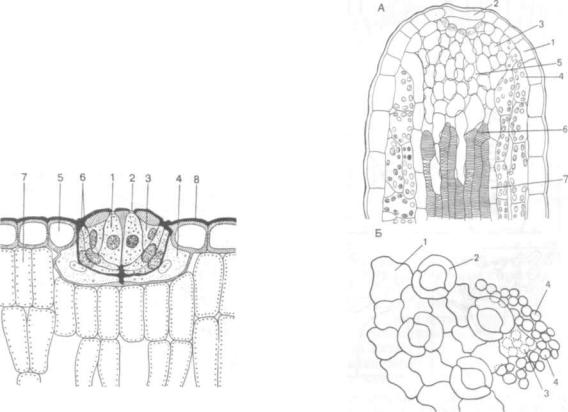
Вместилища выделений. Встречаются у расте ний разных систематических групп, разнообразны по форме, размерам, происхождению. По проис хождению различают схизогенные и лизигенные вместилища.
Схизогенные вместилища (греч. schizo — рас щепляю, раскалываю) образуются за счет расши рения межклетников в молодых тканях. При этом клетки, прилегающие к вместилищу, становятся эпителиальными, т. е. вырабатывают и выделяют в полость экскреторное вещество. Они представ лены одним слоем крупноядерных клеток, запол ненных густой цитоплазмой. Эпителий изолирует экскрет от живых тканей.
Схизогенные вместилища представлены удли ненными ходами, идущими по всему органу, ино гда переходящими в другие органы, округлыми или продолговатыми железками. Каналообразные ходы чаще встречаются в стеблях, корнях, реже — в листьях и содержат смолы, эфирные масла, слизи, камеди. Они встречаются у разнообразных
157
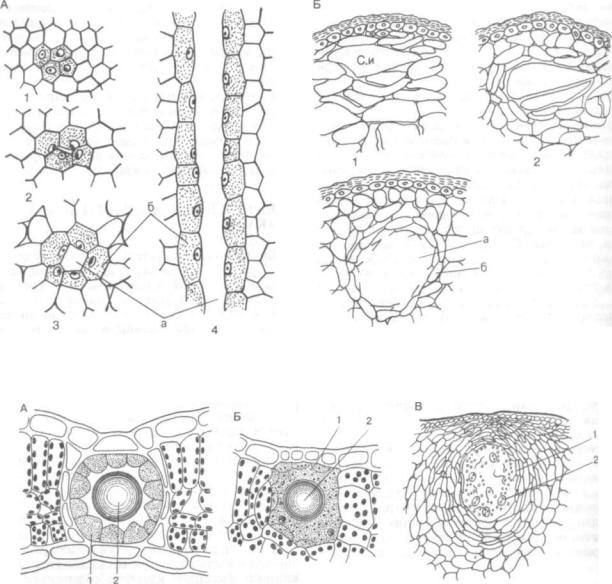
Рис. 123. Смоляные ходы:
Л — ели обыкновенной (Ксеа abies): 1—3 — на поперечных срезах; 4 — в продольном разрезе; Б — в коре побега можжевельника обыкновенного (Juniperus communis): 1—3 — последовательные этапы формирования лизигенного вместилища; а — полость канала; б — эпителий, Си — смоля ной идиобласт
Рис. 124. Типы вместилищ эфирных масел на поперечном срезе:
А — схизогенное вместилище в листе зверобоя (Hypericum perforatum); Б — лизигенное вместилище в листе ясенца (Dietamnus fraxinella); В — лизигенное вместилище в кожуре плода мандарина (Citrum unshiu); 1 — эпителий; 2 — эфирное масло
групп растений — от водорослей (например, ла минариевых) до покрытосеменных. Особенно характерны для хвойных (роды сем. сосновых, Pinaceae — сосна, ель, лиственница, пихта, кедр и др.) (рис. 123, А).
В типичных случаях схизогенные смоляные ходы представлены сложной сетью продольных и поперечных каналов, пронизывающих органы хвойных растений.
Различные филогенетические группы расте ний накапливают в схизогенных вместилищах разные экскреторные вещества: виды миртовых, сельдерейных — летучие терпены; аралиевые — вязкие бальзамы; стеркулиевые, мараттиевые — камеди, слизи и т. д. (рис. 124).
Лизигенные вместилища (греч. lysis — раство рение) возникают в результате растворения со седних клеточных стенок накапливающимися выделениями. В результате возникают ходы или железы внутренней секреции, окруженные слоем эпителиальных клеток.
У можжевельника смоляной ход образуется из смоляного идиобласта (см. рис. 123, Б, I). Коли чество смоляных выделений постепенно реличивается. Под влиянием их нарастания стенки кле ток подвергаются лизису и формируется шарооб разная смоляная железка. Лизигенные вместили ща характерны для цитрусовых (рис. 124, Б), многих зонтичных.
Между схизогенными и лизигенными вмести лищами существуют различные переходные фор мы, в некоторых случаях они образуются одно временно и лизигенно, и схизогенно. Железы внутренней секреции смешанного происхождения хорошо выражены в листьях зверобоя, лавра бла городного, камфорного дерева и др.
Иногда внутренние выделительные ткани представлены цепочками клеток в виде продоль ных рядов, сопровождающих проводящие пучки (фасоль, листья бука, ароидные). В них могут накапливаться дубильные вещества, рафиды (у лилейных).
158
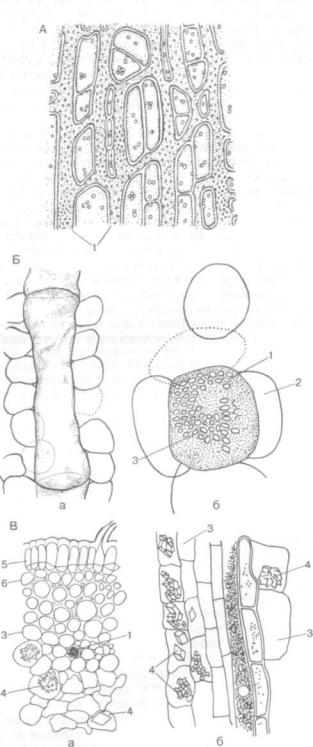
- ' » ' — I —
Млечники (млечные трубки). Представлены отдельными клетками и продольными цепочками слившихся клеток, содержащих в вакуолях млеч ный сок. Как элементы выделительных тканей покрытосеменных растений млечники встречают ся почти у 12 500 видов (в основном сем. моло чайные, колокольчиковые, астровые, маковые), представляющих разные жизненные формы — деревья, травы, кустарники, лианы.
Млечники — живые клетки. Вполне сформи ровавшиеся и функционирующие млечники, изу ченные с помощью электронного микроскопа, содержат постенный слой цитоплазмы с много численными ядрами, крупную вакуоль с млечным соком. Оболочка млечников отличается высокой эластичностью, состоит в основном из клетчатки, не подвергается одревеснению, поры малочис ленные и слабо заметные.
ствуют разные мнения о значении млечников и млечного сока. Некоторые авторы выделяют не сколько основных функций млечников: экскре торную, проводящую, запасающую. Проводящая функция осуществляется при транспортировке запасных продуктов от места их синтеза к ме стам потребления. В таком случае флоэмная часть пучка развита слабо, а млечники часто
Млечный сок представляет собой жидкость молочного цвета (отсюда название), иногда ок рашен в другие цвета (у чистотела — яркооранжевый, у мака — желтый, у конопли — жел товато-коричневый, у шелковицы, олеандра — прозрачный). В мякоти плодовых тел некоторых грибов имеются толстостенные гифы (аналог млечников) с млечным соком различного цвета.
По физическому состоянию млечный сок — это эмульсия: различные вещества (терпены — смолы, каучук, алкалоиды, таннины, белки) на капливаются в виде гидрофобных капелек, взве шенных в водянистом клеточном соке. Он может содержать также амилопласты.
По происхождению и структуре все млечники можно разделить на две большие группы — чле нистые и нечленистые.
Членистые, или сложные, млечники возникают из многих отдельных млечных клеток, которые в местах соприкосновения друг с другом растворя ют оболочки, а их протопласты и вакуоли слива ются в единую разветвленную систему (рис. 125, А, Б). Они найдены у маковых, лобелиевых, колокольчиковых, астровых (роды цикорий, латук). Членистые млечники чаще всего бывают с ана стомозами — соединениями (рис. 125, А), но мо гут быть и без них, как у некоторых лилейных, например у лука (рис. 125, Б). Местоположение членистых млечников разнообразно: у гевеи они сосредоточены во флоэме, в листьях цикория пронизывают весь орган.
Нечленистые, или простые, млечники представ ляют одну гигантскую клетку, которая, возникнув у зародыша, растет, распространяясь во все орга ны растений. Важная особенность этой клетки состоит в том, что ее интрузивный рост и деление ядра не сопровождаются заложением перегоро док. Образуются многоядерные клетки, не сли вающиеся в единую систему. В одних случаях они не ветвятся и сохраняют более или менее цилиндрическую форму (крапива, конопля, оле андр; рис. 125, В), в других — сильно ветвятся и пронизывают все органы (шелковица, молочай).
Несмотря на то что млечники изучаются дав но, сведения об их морфологии, физико-хими ческом составе, физиологических функциях еще не достаточны, а нередко противоречивы. Суще
содержат перфорационные пластинки (рис. 125,
Б, б).
О выделительной и запасающей функциях млечников можно судить в какой-то степени по составу млечного сока. У многих растений млеч ный сок содержит не только конечные продукты обмена веществ (таннины, алкалоиды, органиче ские кислоты, соли кальция и калия), но и по бочные вещества метаболизма (терпены, каучук, смолы и др.). Наряду с этим в млечном соке на ходятся типичные запасные продукты, иногда ферменты. Видимо, у определенных групп расте ний на разных этапах развития млечники выпол няют разнообразную роль. Это подтверждается разным химическим составом млечного сока у растений и изменением его количества на разных фазах онтогенеза.
Млечники, содержащие горькие и ядовитые вещества, защищают растения от поедания жи вотными, млечный сок способствует заживлению ран, так как поверх их свертываются каучукоподобные вещества. Запасные продукты в виде крахмала содержат млечники молочая, запасные белки находятся в млечном соке фикуса, саха ра—у цикория, жиры и ферменты — у дынного дерева, каучук — у каучукового дерева, берескле та и т. д. Отмечают важную роль млечников в регуляции водного и кислородного балансов рас тений, поскольку млечный сок легко поглощает воду и кислород из соседних тканей.
Выяснение функционального и эволюционно го значения млечников осложняется тем, что они
не являются обязательным структурным компо нентом всех высших растений.
Ценность многих растений, обладающих млечным соком, давно известна человеку. Расте ния, млечный сок которых богат сахарами, крах малом, белком, служат хорошими кормовыми травами. Многие тропические растения с млеч ным соком используют как наркотические. Упот ребляют в пищу млечный сок коровьего дерева (Южная Америка), по составу сходный с коровь им молоком.
Особую ценность представляют растениякаучуконосы, в млечном соке которых содержится значительное количество каучука и гуттаперчи. Млечный сок этих растений обычно называют латексом (многие авторы используют это назва ние для млечного сока вообще). Широко извест ное каучуковое дерево (Hevea brasiliensis) — глав ный источник каучука на мировом рынке. Такие растения, как эвкоммия (Китай), гваюла (Мексика), также распространенные каучуконо сы, культивируемые во многих странах мира.
В 30-е годы советские ботаники провели серь езную работу по отысканию дикорастущих кау чуконосных растений. Были найдены травяни стые каучуконосы — кок-сагыз, тау-сагыз, крымсагыз, произрастающие в горах Кавказа, Крыма, Казахстана, введенные затем в культуру. Одно временно химики развернули работу по созданию синтетического каучука. В настоящее время про изводство синтетического каучука свело к мини муму выращивание каучуконосов.

Глава 4
Вегетативные
органы
растений
-<*>
ОБЩЕЕ ПОНЯТИЕ О ВЕГЕТАТИВНЫХ ОРГАНАХ
Вегетативные органы (лат. vegetatio — произ растание, рост), представляющие вегетативное те ло растения, выполняют основные функции пи тания и обмена веществ с внешней средой. На основе строения вегетативного тела все растения подразделяются на две большие группы: низшие и высшие. Низшие растения многоклеточные (водоросли), а также грибы более примитивны и однообразны по строению в связи с водным обра зом жизни или гетеротрофизмом. Тело их пред ставляет однородное слоевище, или таллом, и ли бо совсем не дифференцировано на органы и ткани (нитчатые, некоторые пластинчатые водо росли, мицелии грибов), либо расчленено на специализированные части, которые внешне сходны с органами высших растений, но не име ют сложного тканевого строения (многие круп ные зеленые и бурые водоросли). У высших рас тений вегетативное тело дифференцировано на органы, а органы — на ткани. В эволюции веге тативные органы возникли в результате усложне ния тела растений при выходе на сушу и освое ния воздушной и почвенной сред. Контрастные условия окружающей среды были основной дви жущей силой адаптивной специализации расте ний.
Основные вегетативные органы растений —
листостебельные побеги (обеспечивают фотосин тез) и корни (обеспечивают водоснабжение и ми неральное питание).
Основные органы тела высших растений за ложены уже в зародыше семян (или спор), кото рые при прорастании формируют проросток, со держащий корень, стебель, листья специфиче ской морфоструктуры и тканевого строения (рис.
126). Корни образуют корневую систему, а стебель и листья — побег. У большинства высших расте ний побег располагается над почвой, а корневая система — в почве. Развитие проростка приводит к формированию взрослого растения с типичной структурой и функцией каждого вегетативного органа.
Большинство растений характеризуется осо бенностями роста и формирования: типичные растения растут и надстраивают свое тело в тече ние всей жизни; параллельно нарастанию новых участков тела идет отмирание старых. Рост и формообразование происходят ритмично, поэто му к уже имеющимся участкам тела периодиче ски подстраиваются новые, и тело растения ока зывается состоящим из однотипных участков, повторяющих друг друга. Повторяемость строе ния тела во времени и в пространстве (метамерность) свойственна всем многоклеточ ным растениям — от водорослей до цветковых.
У большинства растений в течение жизни уве личивается число меристематических зон, каждая из которых дает начало новому участку тела, сходному с предыдущим и соседним. Происходит ветвление — процесс, характерный для расти тельных организмов, в результате чего большин ство растений представляет собой разветвленную систему (или системы) из однотипно построен ных частей.
Повторяемость строения вдоль продольной оси тела и разветвленность — важнейшие особенности внешнего строения тела растения.
Одна из общих закономерностей организации структуры вегетативных органов — симметрия. Обычно она наблюдается в характере расположе-
11 Зак. 1851 |
161 |