

Морфология растений / Бавтуто Г.А., Ерёмин В.М. Ботаника. Морфология и анатомия растений
.pdfктеток с горизонтальными конечными стенками; более примитивные — из клеток прозенхимной формы с вытянутыми заостренными концами, их боковые стенки незаметно переходят в конечные (рис. 102, 3—6). Ситовидные трубки обычно тя нутся вдоль продольной оси органа, но есть и поперечно идущие, соединяющие группы прово дящих тканей.
Оболочки ситовидных трубок целлюлозные, лишь к концу вегетации некоторые ситовидные трубки одревесневают. В полостях ситовидных трубок долго сохраняется живой протопласт в виде пристенного слоя, ядро в зрелых элементах отсутствует, живут клетки-членики, как правило, одну вегетацию. Около ситовидных трубок име ются сопровождающие клетки (клетки-спут ницы), структурно и функционально связанные с ними.
Гистогенез ситовидной трубки. В онтогенети I ческом развитии ситовидной трубки выделяется ряд специфических особенностей. Клетка мери стемы, дающая начало ситовидным элементам, вначале делится продольно. Две дочерние клетки в дальнейшем развиваются разными путями, но сохраняют многочисленные плазматические свя зи. Одна из клеток (большего размера) диффе ренцируется в членик ситовидной трубки, дру гая—в клетку-спутницу. Иногда исходная клетка делится 2—3 раза и образуются две-три клеткиспутницы при одной ситовидной трубке (рис. 103, А, Б). Наблюдается и поперечное деление клетки-спутницы с последующим образованием нескольких сопровождающих ктеток, располо женных продольным рядом вдоль ситовидной
трубки.
Первый этап развития ситовидных элементов характеризуется ростом, для него свойственны все особенности растущих клеток. Одновременно с ростом ситовидного элемента происходят спе
цифические изменения в структуре клеточных стенок. Продольные (боковые) стенки у многих видов получают так называемые перламутровые утолщения. Оболочки с таким утолщением дают положительную реакцию на целлюлозу и пекти ны. Перламутровая оболочка в некоторых случаях настолько утолщается, что закрывает просвет клетки. В поперечных стенках в это же время на чинается процесс, который в дальнейшем приво дит к формированию ситовидных пластинок. На блюдается интенсивное развитие органоидов, от ветственных за синтез и секрецию веществ клеточной оболочки — аппарата Гольджи и эндоплазматического ретикулума, в значительном ко личестве встречаются митохондрии и пластиды. Характерным компонентом структуры созреваю щих ситовидных элементов является «флоэмный белок» («Ф-белок»). По химической природе он близок к сократительным белкам микротрубочек и микрофиламентов цитоплазмы (рис. 103, Б).
Все последующие после завершения роста клеток этапы формирования ситовидных элемен тов носят деградационныи характер: цитоплазма и нуклеоплазма сокращаются в объеме, происхо дит распад тонопласта и эндоплазматического ретикулума, затем ядра. Клетка теряет рибосомы, диктиосомы, частично сохраняются митохонд рии, пластиды, причем в сильно измененном ви де. Одновременно формируются ситовидные поля (на боковых стенках) и ситовидные пластинки (на поперечных).
Ситовидная пластинка в общих чертах — это группа перфораций, выстланных плазмолеммой, расположенных на общей для двух члеников по перечной стенке. Перфорации ситовидной пла стинки — результат дифференциации плазмодесм. Развитие сита начинается с дифференциа ции окаймляющей поверхности плазмодесм. Вдоль этой поверхности происходит отложение
Рис. 103. Гистогенез ситовидных элементов (схемат.):
А — меристематические клетки; Б — образование клеток, дающих начало ситовидной трубке и клеткам-шутницам; В — ситовидный элемент с перламутровой оболочкой и Ф-белковым телом; Г — ядро в ситовидном элементе дегенерирует, тонопласт частично разрушен, Ф-белок рассеян по клетке, в будущих ситовидных пластинках образовались срединные полости; Д — зрелый ситовидный элемент, в ситовидных пластинках сфор мировались отверстия, которые выстланы каллозой и содержат Ф-белок; Е— образование мозолистого тела (12); 1 — ситовидная трубка; 2 — клет ки-спутницы; 3 — перламутровая оболочка; 4 — ядро; 5 — Ф-белок; 6 — тонопласт, 7 — плазмодесмы; 8 — пластида; 9 — срединная полость; 10 — отверстие ситовидной пластинки; 11 — каллоза
142

аморфного вещества, в составе которого преобла дает каллоза. Основной материал оболочки при этом подвергается лизису, освобождая место для каллозных отложений. Конечный этап формиро вания отверстий ситовидной пластинки — рас ширение плазмодесменных каналов за счет уп лотнения (или удаления) каллозных образований (рис. 103, Г). Ситовидные поля на боковых стен ках ситовидных трубок имеют более разнообраз ную структуру, составляющие их элементы могут сохранять различную степень сходства с плазмодесмами.
В зрелых ситовидных элементах плазмалемма окружает протопласт, содержащий небольшое число митохондрий, пластид и агранулярный эндоплазматический ретикулум. Тонопласт разру шен, ядро отсутствует. Цитоплазма теряет полу проницаемость и становится вполне проницае мой для растворов органических и неоргани ческих веществ. Ситовидные пластинки имеют перфорации, выстланные плазмалеммой и запол ненные каллозой и фибриллами «Ф-белка», кото рые ориентированы продольно. На этом форми рование ситовидной трубки в основном заверша ется.
У двудольных растений ситовидные трубки функционируют чаще 1—2 года, затем камбий образует новые элементы. Известны случаи, когда этот срок превышает 2—3 года, а у некоторых растений, лишенных камбиального прироста, — и более длительное время, например у винограда, липы и других древесно-кустарниковых пород. У некоторых папоротников ситовидные элементы «работают* до 5—10 лет, а у некоторых пальм да же десятки лет. По мере старения ситовидной трубки, чаще к осени, резко увеличивается коли чество каллозы на ситовидных полях. В результа те поровые поля, которые весной выглядели как углубление в оболочке, вначале выравниваются, а затем приподнимаются над ней. Таким образом, к осени ситовидное поле бывает покрыто плотным слоем каллозы (мозолистое тело). Функциониро вание ситовидных трубок прекращается, так как они становятся непроницаемыми для пластиче ских масс. Впоследствии отмершие трубки сплю щиваются давящими на них соседними живыми клетками. В отдельных случаях (например, у ви нограда) весной каллоза рассасывается и си товидная трубка вновь начинает функциониро вать.
Ситовидные трубки имеют общий план строе ния и различаются у разных растений лишь в де талях. Так, длина ситовидной трубки картофеля равна 138—140 мкм, в листовом черешке настур ции около 240 мкм, самая длинная ситовидная трубка достигает 550 мкм. Длина отдельных чле ников колеблется в пределах 150—300 мкм, попе речник — 20—30 мкм. Поперечники канальцев
• ситовидных полей — около 0,5—0,7 мкм.
t« Клетки-спутницы, или сопровождающие клет- $ ки. Образуются из одной материнской клетки с ситовидной трубкой (рис. 104) и функционально связаны с ней. Это паренхимные элементы флоэ мы, обеспечивающие регуляцию передвижения веществ по флоэме, они связаны с ситовидными
Рис. 104. Строение ситовидной трубки (Л) и клетки-спутницы
(Б) (схемат.):
1 — цитоплазма; 2 — плазмалемма; 3 — плазмодесмы; 4 — ситовидная пла стинка; 5 — каллоза в порах ситовидной пластинки; 6 — Ф-белок; 7 — ядро; 8 — митохондрия; Р — пластиды; 10 — вакуоль; 11 — аппарат Гольджи; 12 — рибосомы; 13 — эндоплазматическии ретикулум; В — поперечный срез
элементами плазмодесмами. Протопласты клетокспутниц отличаются метаболической активно стью: ядро и ядрышко крупные, множество хлоропластов, крупных митохондрий, рибосом, име ется эндоплазматическии ретикулум.
Клетки вакуолизированы, часто содержат мно го мелких вакуолей. Особенностью клеточных стенок является не только наличие многочислен ных ветвящихся плазмодесм (например, в листе ясеня их до 24 000 на клетку), но и обилие в них инвагинаций («лабиринтов»), выстланных плаз малеммой, что значительно увеличивает поверх ность (см. рис. 104). В целом клетки-спутницы по своей ультраструктуре сходны с секреторными клетками, и их способность вьщелять сахар в проводящие элементы против градиента концен трации позволяет предполагать, что они выпол няют секреторную функцию.
Активный метаболизм в клетках-спутницах, тесная связь их с ситовидными трубками, в зна чительной степени утративших признаки само стоятельных живых систем, указывают на актив ную роль сопровождающих клеток во флоэмном транспорте. Предполагают, что в клетках-
143
спутницах интенсивно вырабатываются различ ные ферменты, которые передаются в ситовидные трубки. Следовательно, флоэму надо рассматри вать как единую транспортную систему, состоя щую из проводящих элементов и смежных с ни ми клеток. Возможно, что клетки-спутницы слу жат также источником энергии для транспорта веществ в ситовидных трубках.
Основными веществами флоэмного тока слу жат сахара, главным образом сахароза. Транспор тироваться по ситовидным трубкам могут и дру гие сахара — рафиноза, стахиоза (у ясеня), сорбит у яблони, спирты. Во флоэмном соке обна ружены также азотистые вещества (белки, ами нокислоты — не более 0,5 %), органические кислоты, витамины, фитогормоны (ауксин, гиббереллин и др.), неорганические соли (до 1-3 %).
Передвижение пластических веществ (асси милятов) по ситовидным трубкам происходит со скоростью 50—100 см/ч и включает в себя три взаимосвязанных процесса — загрузку флоэмы, транспорт ассимилятов по ситовидным элементам и разгрузку флоэмы. Особая роль здесь принад лежит, по-видимому, Ф-белку, обладающему со кратительными свойствами, а также живому со держимому ситовидных элементов и особенно клеток-спутниц, которые активно, т. е. с затратой энергии, участвуют в передвижении ассимилятов. С этим предположением согласуется тот факт, что передвижение ассимилятов требует интен сивного дыхания клеток флоэмы: если дыхание затруднено, то передвижение останавливается.
ц Эволюция ситовидных элементов. Элементы, сходные с флоэмой, уже имелись у некоторых водорослей, т. е. в эволюционном плане флоэма значительно старше ксилемы. Более того, можно сказать, что основной физиологический признак клеток флоэмы — способность проводить раство ренные органические вещества — в той или иной степени вообще присущ любой живой клетке, и, следовательно, механизм, обеспечивающий эту способность, возник на самых первых этапах раз вития жизни на Земле. При специализации от дельных групп клеток высших растений на про ведение органических растворов была выработана определенная морфоструктура флоэмнои ткани, но суть оставалась той же.
Рассматривая эволюцию флоэмы, необходимо отметить, что тонкие целлюлозные оболочки ее клеток плохо сохраняются в ископаемом состоя нии в отличие от клеток ксилемы с одревеснев шими стенками. Поэтому сведения о последова тельных этапах развития элементов флоэмы более отрывочны.
Предполагают, что у самых первых наземных растений — риниофитов — клетки флоэмы отли чались от фотосинтезирующих или запасающих паренхимных клеток вытянутой вдоль оси фор мой. Однако даже такое структурно упрощенное растение, как псилот, уже имеет на стенках про водящих элементов мелкие, своеобразно устроен ные отверстия (ситовидное поле, или ситечко).
В процессе эволюции возникли высокоспе циализированные поры (расширенные плазмо-
десмы), которые встречаются у всех высших рас тений, кроме риниофитов, но порядок их распо ложения различен.
У всех споровых растений (хвощевидных, плауновидных, папоротниковидных) и у голосе менных небольшие ситовидные поля рассеяны по всей длине оболочки ситовидного элемента (ситовидные клетки; см. рис. 102, 1, 2). У цветко вых они образуют ситовидные пластинки в сито видных трубках. Следует отметить, что в сито видных клетках часто сохраняется ядро, правда, в полуразрушенном состоянии (так называемые некротические ядра), тогда как ситовидные труб ки лишены ядра.
Клетки-спутницы появились на очень высо ком уровне эволюционного развития флоэмы. У высших споровых флоэма не содержит специали зированных сопровождающих клеток. Клетки Страсбургера (альбуминовые) голосеменных рас тений, иногда встречающиеся рядом с ситовид ными клетками, в структурном отношении пред ставляют нечто среднее между клеткамиспутницами и клетками флоэмнои паренхимы покрытосеменных. У цветковых растений клеткиспутницы связаны с ситовидной трубкой единым происхождением, структурой и функцией. У мно гих примитивных покрытосеменных (например, австробайлея пятнистая — Austrobaileya maculata) они отсутствуют.
У более примитивных семейств цветковых растений ситовидные трубки во многом сходны с ситовидными элементами голосеменных: они длинные, боковые стенки их незначительно ско шены, ситовидные пластинки состоят из 10 и бо лее отдельных ситовидных полей, располагаю щихся на боковых стенках (см. рис. 102, 3, За). Промежуточный тип ситовидных трубок имеет короткие ситовидные элементы, ситовидные пла стинки у которых скошены, с небольшим числом (до 10) ситовидных полей (см. 102, 4, 4а).
Наиболее совершенные ситовидные трубки двудольных имеют относительно короткие клет ки с поперечной стенкой, на которой располага ется ситовидная пластинка, где уже невозможно выделить отдельные ситовидные поля (см. рис. 102, 5, 5а, 6, 6а).
Клетки-спутницы претерпели также некото рую эволюцию. Наиболее примитивной является клетка-спутница, возникшая в результате одного митотического деления вместе с клеткой — буду щей ситовидной трубкой. Последующая эволю ция привела к появлению нескольких клетокспутниц при продольном и поперечном делении исходной клетки. Таким образом, можно гово рить о специализации ситовидных элементов в ходе филогенеза.
Лубяная паренхима. Это постоянный компо нент флоэмы. В клетках лубяной паренхимы ак тивно протекают обменные реакции и накапли ваются различные эргастические вещества - крахмал, жиры, различные органические соеди нения, а также таннины, смолы, кристаллы. Клетки первичной флоэмнои паренхимы имеют продолговатую форму и располагаются парал лельно ситовидным трубкам. Связь их осуществ-
144
ляется посредством простых пор, которые собра ны группами и со стороны ситовидных трубок напоминают ситовидные пластинки. Оболочки паренхимных клеток целлюлозные, тонкие, фи зиологическая активность высокая, расположение среди ситовидных элементов беспорядочное.
Вторичная флоэмная паренхима подразделяет ся на две системы: вертикальную и горизонталь ную. В вертикальной системе лубяная паренхима располагается вместе с ситовидными и механиче скими элементами, горизонтальная система пред ставляет паренхиму сердцевинных лучей. Такая структура особенно характерна для древесных растений.
Кристаллообразующая паренхима флоэмы ча ще всего имеет лигнифицированные оболочки со вторичными утолщениями. Паренхимные клетки, расположенные рядом с ситовидными элемента ми, могут участвовать в загрузке и разгрузке про водящих элементов, часто они дифференцируют ся в передаточные клетки с внутренними вырос тами оболочки. Ряд авторов предполагает, что между паренхимными клетками и ситовидными элементами существует онтогенетическое родст во. У многих двудольных членики ситовидных трубок и некоторые паренхимные клетки проис ходят от общих флоэмных инициалей и отмирают вместе с ситовидными элементами (К. Эсау, 1955). Этот тип флоэмной паренхимы занимает промежуточное положение между клеткамиспутницами и запасающей паренхимой.
Склеренхимные элементы. Представлены дре весинными волокнами и склереидами.
Волокна относятся к обычным компонентам первичной и вторичной флоэмы. В первичной флоэме волокна встречаются только в перифери ческой части ткани, во вторичной — распределе ны по всей ткани среди других клеток осевой системы. В зрелом состоянии волокна могут быть живыми или мертвыми, одревесневшими или неодревесневшими. Живые волокна выполняют функцию запаса. У многих видов первичные и вторичные лубяные волокна используются как источники промышленного волокна.
Во флоэме часто встречаются склереиды. Они располагаются в комбинации с волокнами либо отдельной группой и в осевой, и в лучевой сис темах вторичной флоэмы. Как правило, склереи ды образуются в более старых участках флоэмы вследствие склерификации паренхимных клеток. Нередко склерификации предшествует интрузив ный рост клеток, формируются нитчатые и раз ветвленные склереиды. Длинные и тонкие скле реиды напоминают волокна и часто называются волокнистыми склереидами.
Онтогенез флоэмы. Схематично его можно представить следующим образом. Как и первич ная ксилема, первичная флоэма дифференцирует ся на протофлоэму и метафлоэму. Протофлоэма возникает из прокамбиальных клеток в период интенсивного роста органа в длину. Это узенькие клетки, превращающиеся в ситовидные трубки. Клетки-спутницы в протофлоэме отсутствуют, а ситовидные трубки быстро теряют ядро и не мо гут расти.
Метафлоэма возникает обычно в момент за вершения роста органа в длину и представлена ситовидными клетками и клетками-спутницами. Быстро развивающаяся метафлоэма сплющивает членики протофлоэмы, которые у двудольных часто превращаются в лубяные волокна.
У однодольных протофлоэма обычно не со держит лубяных волокон и резко отграничивается от участков метафлоэмы. У двудольных переходы между прото- и метафлоэмой незаметны. У одно дольных метафлоэма сохраняется как активная проводящая ткань, так как вторичная флоэма, в связи с отсутствием камбия, не образуется. У дву дольных камбий формирует вторичную флоэму. Подробнее первичная и вторичная флоэма рас сматривается в темах «Стебель» и «Корень».
ПРОВОДЯЩИЕ ПУЧКИ
Проводящие элементы в комплексе с парен химными и механическими элементами образуют
втеле растения тяжи, которые называют проводя щими пучками.
Вмолодых органах большинства растений проводящие пучки идут раздельно. На более поздних стадиях развития органов у двудольных и голосеменных растений проводящие пучки сли ваются, образуя сплошной цилиндр, состоящий из тканей древесины и луба, так называемые слои проводящих тканей. У однодольных пучковая структура сохраняется на всех стадиях развития органа.
Система проводящих пучков возникла и со вершенствовалась в процессе приспособления растений к наземным условиям существования. Она пронизывает все органы растений, объединяя их в одно целое и обеспечивая в растении еди ный обменный процесс со множеством разнооб разных отправлений. Пучки хорошо видны в ли стьях в виде сети жилок, а также в сочных стеб лях, например у недотроги. Проводящие пучки образуют сложную сеть не только в вегетативных, но и в генеративных органах, особенно в плодах.
Образование пучков. Прокамбий. Камбий.
Формирование проводящих пучков осуществля ется на ранних стадиях развития органов расте ния за счет деятельности специальной меристематической ткани — прокамбия (лат. pro — перед, раньше и позднелат. cambium — обмен, смена). Прокамбий дифференцируется из промеристемы
вапексе побега в области формирования листо вых зачатков (в корне представлен колонкой в центре органа). Клетки сильно вытянутые, тонко стенные, слабовакуолизированные, возникают в стебле в виде небольших участков, расположен ных кольцом. Отличаются от окружающих клеток густым содержимым и физиологической активно стью (рис. 105). Прокамбий откладывает кнаружи органа элементы флоэмы, внутрь — элементы ксилемы.
Первыми в прокамбии дифференцируются элементы протофлоэмы и протоксилемы; вторая ступень дифференциации — метафлоэма и метаксилема. Образовавшиеся из прокамбия прово-
10 Зак. 1851 |
145 |

Рис. 105. Разрез через верхушку стебля льна (Linum usitatissimum) с тяжем прокамбиальных хлеток (I)
дящие пучки, состоящие из флоэмы и ксилемы, называются первичными. В результате работы про камбия формируются два типа проводящих пуч ков. Если все клетки прокамбия дифференциру ются в ткани проводящего пучка, то такой пучок не способен к дальнейшему росту и называется закрытым. Если же между флоэмой и ксилемой остается прокамбиальный слой, дающий начало камбию, пучок продолжает расти благодаря обра зованию новых элементов флоэмы и ксилемы и называется открытым. Закрытые пучки наблю даются обычно у однодольных растений и очень редко у двудольных; открытые — характерны для двудольных и хвойных растений, стебли и корни которых способны к утолщению (рис. 106).
Камбий как меристематическая ткань несколь ко отличается от прокамбия. Он содержит два типа клеток: удлиненные с заостренными конца ми — веретеновидные инициали и почти изодиаметрические, сравнительно небольшие клетки —
лучевые инициали (рис 107). Веретеновидные ини циали дают начало всем элементам проводящих тканей, которые расположены вдоль продольной оси органа (продольные или осевые элементы ксилемы и флоэмы — трахеиды, сосуды, сито видные элементы, волокна, паренхима). Лучевые инициали дают начало клеткам лучей (по перечная, или лучевая, система ксилемы и фло эмы). Плоские широкие стороны каждой клетки камбия обращены внутрь (к ксилеме) и наружу (к флоэме). Остальными стенками клетка смыкается с соседними клетками камбия.
Камбий может быть ярусным и неярусным. В ярусном камбии веретеновидные инициали бо лее короткие и в меньшей степени перекрывают друг друга, чем в неярусном (рис. 108). Ярусный камбий дает начало ярусной древесине, а из не ярусного камбия возникает неярусная древесина.
Камбиальные инициали, делясь периклинально в тангенциальной плоскости (т. е. параллельно плоским сторонам), образуют элементы вторич ной ксилемы и флоэмы. В период наибольшей ак тивности камбия деление клеток происходит бы стро, и к моменту появления новых клеток ранее отложенные еще остаются меристематическими. В результате формируется широкая зона недиф ференцированных клеток, которую называют камбиальной зоной. В ней только один слой пред ставлен делящимся камбием, остальные слои - так называемые флоэмные и ксилемные инициа ли, или материнские клетки проводящих тканей.
Камбиальные клетки сильно вакуолизированы, несут на оболочках первичные поровые поля с многочисленными плазмодесмами. Радиальные стенки этих клеток утолщены сильнее, чем тан генциальные. Камбиальные клетки в 4 раза чаще превращаются в элементы ксилемы, чем в эле менты флоэмы.
Классификация пучков. Проводящие пучки различаются по происхождению, составу, спо собности к росту и т. д.
По элементарному составу различают четыре группы пучков.
Простые пучки по структуре наиболее прими тивны и состоят из однородных гистологических элементов: а) из одних трахеид, например в ли-
Рис. 106. П р о в о д я щ и е пучки:
А — закрытый пучок стебля кукурузы (Zea mays); £ - открытый пучок стебля лютика ползучего (Ranunculus repens); / — флоэма; // — ксилема; 1 — лубяная па ренхима; 2 — склеренхима; 3 — ситовидные трубки; 4 — клетки-спутницы; 5 — сосуды; 6 — древесинная паренхима; 7—камбий
146

Рис. 107. Камбий в стебле табака (Nicotiana tabacum):
Л — радиальный срез: 1 — лучевая инициаль; 2 — веретеновидная инициаль; 3 — образование межклеточной пластинки в делящейся клетке; 4 — фрагмопласт, Б — схема лучевой (а) и веретеновидной (6) шшциалей: пунктиром показана плос кость периклинального деления, приводящая к образованию производных клеток; В — тангенциальный срез: 5 — апикальный интрузивный рост; 6 — наклонная ра диальная стенка; Г— сечение цилиндрического органа: а — поперечное; б — про дольное радиальное; в — продольное тангенциальное
Рис. 108. Типы камбия:
Л — ярусный у акации (Robinia); Б — неярусный у ореха (Juglans); 1 — веретеновидные инициали; 2 — лучевые инициали
стьях многих растений, особенно вблизи края листовой пластинки, или б) из одних ситовидных трубок, например в цветочных стрелках .гука (Allium сера) и др. Общие пучки — трахеиды, со суды и ситовидные трубки расположены бок о бок. Сложные пучки, помимо проводящих элемен тов, содержат паренхимные элементы. Сосудистоволокнистые пучки, наиболее специализирован ные -и широко распространенные, включают все элементы ксилемы и флоэмы.
По наличию или отсутствию камбия, как уже говорилось, пучки бывают открытые и закрытые. В. Г. Александров (1966) в этой группе пучков выделяет элементарный пучок, который содержит первичные анатомические элементы, образован ные прокамбием, и вторичные элементы, образо ванные продолжающим функционировать в пучке камбием.
Расположение ксилемы и флоэмы может быть различным, и в связи с этим выделяют следую щие типы проводящих пучков.
Коллатеральным (лат. con — вместе, с и lateralis — боковой) или бокобочным называют пучок, когда флоэма и ксилема располагаются бок о бок, т. е. на одном радиусе (рис. 109, 1; рис. ПО, А). Наружная часть пучка обычно представ лена флоэмой, внутренняя — ксилемой. Этот тип пучка наиболее распространен и встречается в листьях всех семенных растений, в осевых орга нах всех однодольных и многих травянистых дву дольных. Формально к этому типу можно отнести и строение осевых органов древесных растений, полагая, что весь ствол или корень содержит лишь один пучок.
Биколлатеральный, или дважды бокобочный, пучок — флоэма прилегает к ксилеме с обеих сто рон, один участок флоэмы более мощный — на ружный, другой — слаборазвитый — внутренний (рис. 109, 2; рис. ПО, Б). Эта форма проводящих пучков присуща растениям из сем. тыквенных, пасленовых, колокольчиковых, сложноцветных. Биколлатеральные пучки образуются, по-види-
147
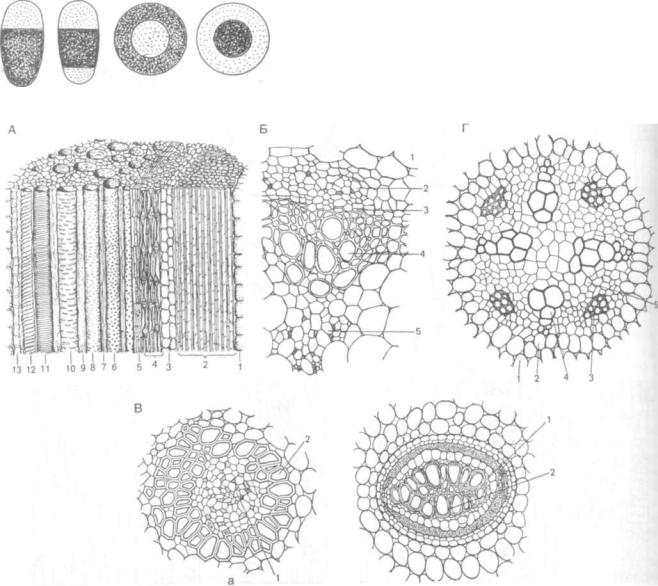
Рис. 109. Схемы типов проводящих пучков:
1 — коллатеральный; 2 — биколлатеральныи; 3 — концентрически! амфивазальный; 4 — концентрический амфикрибральный; 5 — ради альный. Флоэма светлая, ксилема темная
Рис. 110. Типы проводящих пучков:
А — коллатеральный открытый сосудисто-волокнистый проводящий пучок подсолнечника (Helianthus annuus) в продольном разрезе: 1 — основная паренхи ма; 2 — склеренхима; 3 — лубяная паренхима; 4 — ситовидные трубки с клетками-спутницами; 5 — камбий; 6 — пористый сосуд; 7 — древесинная паренхима; S — сетчатый сосуд; 9 — либриформ; 10 — лестничный сосуд; 11, 12 — спиральные сосуды; 13 —• кольчатый сосуд; Б — биколлатеральныи проводящий пучок картофеля (Solanum tuberosum) на поперечном срезе: 1 — основная паренхима; 2 — наружная флоэма; 3 — камбий; 4 — ксилема; 5 — внутренняя флоэма; В — концентрические пучки: а — амфивазальный из корневища касатика (Iris); б — амфикрибральный из корневища орляка (Pteridium aquilinum): / — флоэма; 2 — ксилема; /— радиальный проводящий пучок корня подсолнечника на поперечном срезе: 1 — эндодерма; 2 — перишпсл; 3 — флоэма; 4 — ксилема; 5 — парен хима
мому, в результате слияния двух коллатеральных пучков.
Концентрический пучок встречается относи тельно редко. Различают два варианта: а) амфива зальный, в котором ксилема замкнутым кольцом окружает флоэму (рис. 109, 3); встречается у од нодольных, например в корневище ландыша, касатика (рис. ПО, В, а), во вторичном приросте стебля у древовидных однодольных (например, у драцены), из двудольных — у клещевины; ^ам фикрибральный, в котором флоэма окружает кси лему (рис. 109, 4). Встречается у папоротнико видных, например у орляка (рис. ПО, В, б).
В радишьном пучке участки флоэмы и ксиле мы лежат по разным радиусам, разделены паренхимной тканью (рис. 109, 5). Этот тип пучка ха рактерен для первичного строения корня у дву
дольных растений (рис. ПО, Г). В корне однодольных такие пучки сохраняются до конца жизни. У двудольных при переходе от первичного ко вторичному строению корня радиальное рас положение флоэмы и ксилемы сменяется колла теральным.
У разных растений при формировании пер вичной структуры корня развивается строго опре деленное число лучей ксилемы и флоэмы, при этом число лучей флоэмы равно числу лучей кси лемы. Встречаются однолучевые (монархные), двулучевые (диархные), трехлучевые (триархныё), четырехлучевые (тетрархные, рис. 110,7) и много лучевые (полиархные; греч. arche— начало, проис хождение) радиальные пучки. Наиболее распростра нены пятилучевые {пентархные). Ксилема в центре корня обеспечивает сопротивление на разрыв.
148

Во всех открытых пучках, где из прокамбия дифференцируется камбий, структура динамична и постепенно усложняется. Камбий, расположен ный внутри пучка, называется пучковым. Он представлен или сплошным замкнутым кольцом, или в форме тяжей, отграниченных от основной паренхимы. Работа камбия приводит к разраста нию пучков в радиальной и тангенциальной плоскости. В стебле часто после начала деятель ности пучкового камбия из живых паренхимных клеток, образующих первичные сердцевинные лучи, возникает межпучковый камбий. Участки межпучкового камбия, соединяясь с участками пучкового камбия, образуют камбиальное кольцо (рис. 111).
Рис. 111. Образование камбиального кольца (схемат.): 1 — пучковый камбий; 2 — межпучковый камбий
Межпучковый камбий начинает формировать новые пучки, камбий в пучках — новые элементы ксилемы и флоэмы. Постепенное разрастание пучков нарушает пучковую структуру. В процессе формирования пучков происходит не только но вообразование анатомических элементов, но и их отмирание и удаление.
В ходе развития высших споровых и семенных растений типы проводящих пучков совершенст вовались. Их структура служит показателем уров ня эволюционного развития. Наиболее прими тивен концентрический, а наиболее эволюционнно продвинут коллатеральный пучок открыто го типа.
ПАРЕНХИМНЫЕ ТКАНИ
Паренхима — основная ткань, составляющая большую часть тела растений, внутри которой дифференцируются высокоспециализированные ткани.
Слово «паренхима» происходит от греч. para — рядом и enchyma — налитое. Этот термин отража ет старое представление о паренхиме как о полу жидком веществе, как бы разлитом вокруг тка ней с более плотной структурой.
Несмотря на то что некоторые авторы возра жают против классификации паренхимы как ос новной ткани, это в какой-то мере оправдано как в морфологическом, так и в физиологическом
смысле. Термин «основные ткани», предложен ный немецким ботаником Ю. Саксом, быстро стал общепринятым. Действительно, эти ткани основные не только потому, что составляют ос новную массу всех органов, в которую как бы инкрустированы арматурные и проводящие тка ни. Они являются основными и потому, что в ходе онтогенеза, при развитии из семени проро стка, служат монолитной основой апексов, из которых развиваются разнообразные по структуре органы тела растений. Наконец, в ходе историче ского развития паренхимные ткани составляли основу всех многоклеточных растительных орга низмов и предшествовали другим тканям. У рас тений более высокого эволюционного уровня основные ткани прошли сложный путь диффе ренциации, специализируясь на выполнении соответствующей функции. Эти функции зависят от происхождения, структуры и расположения тканей в теле растений и лежат в основе их клас сификации.
Основные функции паренхимных тканей свя заны с синтезом и запасанием органических ве ществ. Эти ткани — источник и потребитель уг леводов в растении. В вакуолях паренхимных клеток могут накапливаться запасные белки, жиры, антоцианы, таннины и другие вещества. У многих растений сухих местообитаний в па ренхиме запасается вода, у водных растений — газы.
К паренхимным тканям относятся и раз личные типы секреторных клеток, специализи рующихся на синтезе специфических белков, жиров, полисахаридов, терпенов и т. д. Спектр функций может меняться благодаря физио логической пластичности паренхимных кле ток.
Однако даже на высоких уровнях специализа ции основные ткани сохраняют свои характерные черты. Все клетки основной паренхимы живые, изодиаметрической формы. Оболочки тонкие, с простыми порами, реже утолщенные и одревес невшие. В типичных случаях в этих тканях хоро шо развиты межклетники. Клетки основной па ренхимы не теряют способности к делению и могут возвращаться к меристематическому со стоянию, например при заживлении ран, регене рации тканей и органов, образовании придаточ ных корней. Выделяют несколько групп основ ных тканей.
АССИМИЛЯЦИОННЫЕ, ИЛИ ХЛОРОФИЛЛОНОСНЫЕ, ТКАНИ (ХЛОРЕНХИМА)
Главная функция ассимиляционных тканей — фотосинтез. Именно в этих тканях синтезируется основная масса органических веществ и связыва ется энергия, получаемая землей от солнца. Фо тосинтез определяет характер всей биосферы на шей планеты и делает ее пригодной для жизни человека.
Клетки ассимиляционной паренхимы тонко стенные, содержат хлоропласты, которые чаще
149

Рис. 112. Часть поперечного среза через лист белены черной (Hyoscyamus nigei):
1 — эпидермис; 2 — устьице; 3 — палисадная паренхима; 4 — губчатая паренхима; 5 — кристаллические включе ния; б — проводящие пучки в поперечном и продольном сечениях
всего располагаются одним слоем вдоль стенок, не затемняя друг друга.
В разных органах размеры и форма клеток хлоренхимы неодинаковы. Наиболее разнообраз на хлоренхима листьев (рис. 112). Продолговатая форма таких клеток называется столбчатой или палисадной. Хлоренхима, состоящая из округлых клеток с большими межклетниками, называется губчатой паренхимой. Фотосинтетическая актив ность хлоренхимы листа (мезофилла) пропорцио нальна числу находящихся в ее клетках хлоропластов. Так, процентное соотношение хлоропластов в клетках столбчатой и губчатой паренхимы у земляники (Fragaria vesca) равно 86 и 14.
Важное значение в структуре хлоренхимы иг рают межклетники. Они резко увеличивают зеле ный экран листа, т. е. площадь соприкосновения клеток мезофилла с воздушной средой, из возду хоносных полостей в клетки поступает С02 , не обходимый для фотосинтеза, а в них выделяется 02 . Такую поверхность клеток мезофилла назы вают внутренней поверхностью листа; внешней поверхностью служит эпидермис. Внутренняя поверхность листа во много раз превышает внеш нюю (например, у катальпы в возрасте 20 лет это соотношение равно соответственно 390 м2 и 5200 м2). В некоторых случаях внутренняя по верхность хлоренхимы листа увеличивается за счет образования внутренних складок оболочки клеток, как, например, в листьях-иглах сосны, ели. Эта паренхима называется складчатой.
Хлоренхима образуется также в молодых стеб лях, цветках, плодах. Как правило, она залегает непосредственно под эпидермисом, что обеспе чивает хорошее освещение и газообмен в стебле. Просвечивая сквозь прозрачную кожицу, хлорен хима окрашивает в зеленый цвет молодые части растений. Иногда она располагается в глубине стебля вокруг пучков или более поверхностно под механической тканью. В данном случае ее функ ция, вероятно, связана со снабжением внутрен них тканей стебля и в первую очередь живых клеток проводящих пучков кислородом, который образуется в процессе дыхания.
В редких случаях хлоренхима образуется и в корнях, доступных свету (в воздушных, в корнях водных растений).
ЗАПАСАЮЩИЕ ТКАНИ
Вещества, поступающие из внешней среды или синтезированные растением, могут запасай ся. Вещества запаса легко обнаружить в живых клетках любой ткани, однако массовое отложение органических веществ происходит только в спе циализированных запасающих тканях, являю щихся основным типом тканей ряда органов.
Запасные вещества накапливаются в опреде ленных частях растения: у деревьев и кустарни ков — в паренхимных клетках коры, сердцевин ных лучей, древесинной паренхимы стволов и корней, а у молодых побегов — в клетках сердце вины. У многолетних травянистых растений обычно имеются специализированные органы запаса — корнеплоды, луковицы, клубни, корне вища. Накопленные летом запасы органических веществ расходуются весной на образование мо лодых побегов и корней.
В плодах и семенах запасающая паренхима со ставляет основу органов. Запасающая ткань соч ных плодов является одной из самых крупнокле точных, диаметр клеток достигает 1—2 мм. В се менах однодольных органические вещества накапливаются в специализированной ткани — эндосперме: у злаков, например, в нем выделяют ся упорядоченно расположенные клетки алейро нового слоя; в семядолях, являющихся органом запаса многих двудольных, — столбчатая и губча тая паренхима (рис. 113).
Специализация запасающих тканей определя ется в значительной степени составом аккумули руемых продуктов. Вещества накапливаются в растворимом или твердом состоянии. В семенах, которые по мере созревания сильно обезвожива ются, высыхают, откладываются высокомолеку лярные запасные соединения в виде твердых зе рен (белки, крахмал, гемицеллюлозы); жиры представлены гидрофобными липидами. Чаще всего в запасающих тканях семян присутствуют одновременно два (белок и крахмал или белок и жир) или все три типа основных запасных ве ществ (см. рис. 113). В клубнях, корневищах, околоплодниках, уровень оводненности которых лишь незначительно изменяется по мере созрева ния, редко накапливаются белки или липиды.
150

Г" 2
Рис. 113. Участок запасающей ткани семядоли земляного ореха (Arachis hypogaea):
) — капли масла; 2 — алейроновые зерна; 3 — крахмальные зерна; 4 — поры оболочки
Рис. 114. Водоносная паренхима:
А — водоносные клетки в листьях риса (Oryza sativa); Б — клетки коры корня тамуса обыкновенного (Tamus communis), содержащие слизь и воду
Рис. 115. Аэренхима:
А — в стебле ситника (Juncus efiusus); Б — в листе белокрыль ника болотного (Calla palustris)
Запасными продуктами в этих органах обычно являются высокополимерные углеводы (крахмал, инулин, гемицеллюлозы) или водорастворимые сахара (корнеплоды свеклы, моркови, луковицы лука, стебли сахарного тростника, мякоть плодов арбуза, винограда). Иногда в яблоках и в ягодах винограда в качестве промежуточного продукта накапливаются органические кислоты.
Различие химической природы запасных ве ществ приводит к специализации тканей, прояв ляющейся и на уровне ультраструктуры. Так, от ложение крахмала происходит в амилопластах, местом запасания белков и Сахаров служат вакуо ли клетки, липидные капли накапливаются непо средственно в гиалоплазме, а гемицеллюлозы — в клеточной оболочке.
Растения, периодически испытывающие не достаток воды, иногда образуют особые водонос ные запасающие ткани. Чаще всего эти ткани со стоят из крупных тонкостенных паренхимных клеток, которые заполнены смесью слизи и воды (рис. 114). Водоносная паренхима встречается в стеблях и листьях растений-суккулентов (как тусы, агавы, алоэ), у многих растений солончаков (солерос, саксаул), в листьях некоторых злаков (рис. 114, А). Иногда воду накапливают одиноч ные клетки-идиобласты, а также запасающие тка ни луковиц, клубней.
ВОЗДУХОНОСНЫЕ ТКАНИ (АЭРЕНХИМА)
Во всех органах и тканях растений имеются межклетники, осуществляющие газообмен и со общающиеся с внешней средой через отверстия покровных тканей. В процессе жизнедеятельно сти (фотосинтез, дыхание, испарение) растения выделяют в межклетники одни газы и поглощают другие, поэтому газовый состав в межклетниках сильно отличается от атмосферного. Характер циркуляции газов по межклетникам, обеспечи вающий нормальную жизнедеятельность, обу словлен типом растения, условиями обитания. Во многих случаях в растениях образуется ткань с крупными межклетниками и преобладающей функцией газообмена (вентиляции). Такую ткань называют аэренхимой.
151