

Состав некоторых веществ в плазме крови и в моче в процентах
(Cushny, 1926)
|
Плазма крови |
Моча |
Степень концентрации в почках |
Мочевина ............................................ Мочевая кислота .............................. Креатинин ....................................... Натрий ............................................. Хлориды ......................................... Калий ............................................... Кальций ........................................... Магний ............................................ Фосфаты ........................................... Сульфаты ...................................... |
0,03 0,004 0,001 0,3 0,37 0,02 0,008 0,0025 0,009 0,002 |
2 0,05 0,075 0,35 0,6 0,15 0,012 0,006 0,15 0,18 |
60 12 75 1 2 7 2 2 16 90 |
Почки принимают участие также и в метаболизме белков, жиров и углеводов. Это подтверждается нахождением в почечной ткани ферментативных веществ (а- L-аминоксидаза, гликоколдегидрогеназа, трансамина-за и другие пептидазы). До сих пор остается невыясненным, в каком виде реабсорбируется в канальцах фильтруемый белок (около 40—50 г в сутки) — в неизмененном состоянии или же в виде аминокислот, из которых затем образуются сложные амины. По-видимому, фильтрованный белок в клубочках превращается в аминокислоты, из которых затем образуются амины, а частично аммиачные продукты распада аминокислот. В пользу этого говорят исследования Polonovski и Colin (1957), Boulanger и Siserte (1956), которые доказали, что аммиак синтезируется в почке из аминокислот и основных аминов, в частности из глютамина.
Из экстракта почки получены такие ферменты, как фосфатазы, гексоки-назы, поэтому можно предположить, что почка наряду с печенью играет большую роль в процессе гликогенеза. В пользу этого говорят трудности терапии больных сахарным диабетом, у которых имеет место заболевание почек.
Почка принимает участие и в обмене жиров, что доказывается увеличением уровня жиров в плазме при нефротическом синтезе, а также обнаружением в почечных канальцах трибутиразы и фосфоэнтеразы. Разнообразные функции почки теснейшим образом связаны с ферментативными процессами, которые во многом зависят от митохондрий клеток. Митохондрии содержат основной набор ферментов, какие и позволяют осуществлять различные сложные биохимические реакции внутри клетки: окисления, фосфорилирования, образования аденозинтрифосфата и т. д. Кроме этого, митохондрии клеток почки обладают большой способностью накапливать различные вещества. Митохондрии очень лабильны и при различных заболеваниях могут легко повреждаться, вследствие чего возникает нарушение энзимных процессов в почке, что, в свою очередь, ведет к нарушению ее функций и жизнедеятельности организма.
Для выполнения многообразия функций почки требуется большое количество энергии, этим и объясняется большое потребление почками крови (до 1500 л в сутки). Частично энергетические затраты почками компенсируются процессами окисления продуктов распада углеводов, аэробного фосфорилирования аденозинтрифосфата, а также в процессе окисления жирных кислот. Происходящие в почках энергетические процессы теснейшим образом связаны с функциями митохондрий клеток. Так, Wachstein (1956), используя цветные энзимные реакции, доказал, что в собирательных канальцах почки происходят активные процессы реабсорбции и экскреции и что эти отделы нефрона не являются лишь транспортными элементами, как это предполагали раньше.
В последнее время установлено, что почки принимают участие в эрит-ропоэзе. Клинические наблюдения над больными пиелонефритом, страдающими анемией, а также острая эритробластопения при анурии позволили высказать мнение о том, что в почке образуется эритропоэтический фактор. Это положение подтверждается экспериментальными данными Jacobson, Goldwasser, Fried и Plazak (1957). У животных, подвергнутых кровопусканию и нефрэктомии, исчезает из плазмы эритропоэтический фактор. Пока еще не удалось получить в чистом виде эритропоэтический фактор, однако клинические и экспериментальные данные не позволяют сомневаться в его существовании, как и в том, что он образуется почками.
Почки оказывают определенное влияние на свертывающую систему крови. Еще Sahli (1885) установил фибринолитические свойства мочи, а в последующем Astrup и Sterndorff (1952) определили, что фибринолитиче-ская активность мочи обусловлена наличием в ней активатора плазминогена — урокиназы. В настоящее время еще не ясно, где вырабатывается урокиназа. Возможно, что она является продуктом деятельности почечного эпителия. Участие почки в свертывающих и антисвертывающих процессах крови подтверждается клиническими наблюдениями над больными пиелонефритом. Во время операции этих больных, как правило, наблюдается повышенная кровоточивость тканей, степень которой не всегда соответствует лабораторнымданным исследования как свертывающей, так и антисвертывающей системы крови. Этот вопрос требует еще дополнительного изучения.
Особая роль принадлежит почке в регуляции артериального давления, и она может рассматриваться как гормонально-секреторная ее функция. Ruyter в 1925 г. впервые обнаружил в почках млекопитающих нейромио-эпителиальный юкстагломерулярный комплекс, состоящий из периартериальной прокладки вокруг проводящей артериолы. Эти клеточные скопления, иначе называемые polkissen Циммермана (Zimmerman, 1933), были обнаружены и вокруг отводящей артериолы и эпителиальной пластинки (macula densa). Юкстагломерулярный комплекс обнаружен у детей только старше 2 лет. У детей старшего возраста отмечаются переходные формы от эластического типа до эпителиального афибриллярного типа.
В настоящее время принято считать, что юкстагломерулярный комплекс обладает эндокринной функцией и выделяет ренин, какой участвует в регуляции почечного и общего кровообращения (Goormaghtigh, 1945). А. М. Вихерт и Ю. А. Серебровская (1962), применив микрохирургическую технику, доказали, что ренин вырабатывается в участках почки соответственно расположению юкстагломеруляриого комплекса. Ренин впервые был обнаружен в 1898 г. Tigerstedt и Bergman. Они установили, что экстракт почки после его введения в ток крови животного вызывает повышение артериального давления.
Экспериментально-клинические исследования позволили установить, что изменение почечного кровообращения, нарушение пассажа мочи из почки, развитие перинефрита вызывают возникновение артериальной гипертонии. Goldblatt, Hanzal, Lynch, Summerville (1934) путем сужения почечных артерий в эксперименте удалось наблюдать у животных стойкое повышение артериального давления без нарушения экскреторной функции почек. Этим экспериментам предшествовали исследования Я. Я. Стольникова (1880), который воспроизвел у животных нефрит посредством нарушения кровообращения в почках за счет сжимания почечных артерий. Экспериментальные исследования Goldblatt с сотр. (1934) легли в основу учения о так называемой почечной гипертонии и установили роль почек в регуляции артериального тонуса. Развитие артериальной гипертонии в условиях нарушения почечного кровообращения, а именно возникновения ишемии, объясняется повышенной продукцией ренина, которая связана с нарушением дыхания почечных клеток. Катализаторы дыхания — цитохром С и аминоксидаза — препятствуют образованию ренина (Hamburger с сотр., 1965).
У собак с экспериментальной гипертонией юкстагломерулярный комплекс гипертрофируется и содержит увеличенное количество зернистых вакуолизированных клеток (Goormaghtigh, 1945). Hartroft P. и Hartroft W. (1952) установили, что изменения юкстагломерулярного комплекса связаны с приемом хлористого натрия и дезоксикортикостероида. При ограниченном приеме хлористого натрия клетки юкстагломерулярного комплекса мышей становятся более многочисленными и богатыми зернистостью. Эти данные согласуются с данными Dunihue (1949), которому удалось установить, что у адреналэктомированных животных в клетках юкстрагломерулярного комплекса происходят те же изменения. Эти данные позволили прийти к выводу, что кортикостероиды влияют на юкстрагломерулярный комплекс через посредство хлористого натрия (Hartroft P. и Hartroft W.). Tobian (1960, 1962) установил, что увеличение активности юкстагломерулярного комплекса, выражающейся в появлении большого количества гранул в его клетках, наблюдается при нарушении почечной артериальной гемодинамики, что связано с уменьшением объема крови, поступающей в почку. На основании этого возникла гипотеза о влиянии объема крови (плазмы) на функцию юкстагломерулярного комплекса. При увеличении давления в почечных артериях и соответственно в приводящих артериолах, какие располагаются в непосредственной близости к клеткам юкстагломерулярного комплекса, возникает повышенное давление на его клетки, что проявляется угнетением их секреции. И, наоборот, уменьшение давления в приводящих артериолах, уменьшение объема притекающей крови к почкам вызывает уменьшение просвета артериол, что способствует снижению давления на юкстагломерулярные клетки, следствием чего является повышенная их секреция.
Иначе говоря, на основании этой гипотезы можно предполагать, что юкстагломерулярный комплекс выполняет роль волюморецепторов, реагирующих на изменение объема плазмы.
Все эти наблюдения дали возможность заключить, что юкстагломерулярный комплекс своей функцией влияет не только на артериальное давление, но и участвует в регуляции электролитов, в частности натрия. Эти две функции юкстагломерулярного комплекса, как регуляция артериального давления, так и регуляция электролитного баланса, тесно связаны друг с другом.
В настоящее время установлено, что ренин представляет собой псевдоглобулин, являясь протеолитическим, весьма устойчивым к температуре ферментом. Ренин не обладает гипертензивными свойствами, но при контакте с плазмой крови, а именно с а2-глобулиновой фракцией, в которой, по мнению Braun-Medendez с сотр. (1946, 1958), содержится гипертензиноген (ангиотензиноген), образуется гипертензин (ангиотонин). Гипертензин, или ангиотонин, еще не является активным веществом. При воздействии ренина на ангиотензиноген образуется ангиотонин-I (гипертензин-1) и лишь под влиянием энзимной группы плазмы крови последний превращается в ангиотонин-П (гипертензин-П), который обладает мощным вазо-прессивным действием.
Ангиотонин-I представляет собой декапептид, т. е. состоит из 10 аминокислот, а после воздействия на него энзимов плазмы превращается в октопептид, ангиотонин-П. Ангиотонин-П в организме разрушается ангио-тензиназой (гипертензиназой), которая находится в слизистой оболочке кишечника, эритроцитах, почках и других тканях. Ангиотонин-И обладает вазопрессивным действием на артерии и артериолы, но не вызывает сужения капилляров В венозной крови нормальной почки ренин почти не содержится, в то время как в крови почки при ее ишемии содержание этого вещества очень высокое. Однако содержание ренина в крови при почечной гипертонии порой не соответствует цифрам артериального давления. Если в начале развития почечной гипертонии содержание ренина в крови высокое, то при хронической гипертонии количество ренина в крови становится ничтожным. Это наводит на мысль, что поддержание высоких цифр артериального давления при хронической форме почечной гипертонии происходит не за счет большой продукции почкой ренина, а за счет каких-то других факторов. Примером этому могут служить экспериментальные и клинические наблюдения, когда удаление почки, находящейся в условиях ишемии, не приводит к нормализации артериального давления. Это объясняется развитием в контралатеральной почке явлений атеросклероза. Однако, как нам кажется, в этих случаях нельзя забывать роль нейрогенного, в частности нейро-вегетативного, фактора. В самом начале развития почечной гипертонии основная роль принадлежит гуморальному фактору, а после того как она примет хроническую форму, наряду с поражением сосудов контралатеральной почки включается нейро-веге-тативный механизм, который и создает условия для поддержания высоких цифр артериального давления.
При ежедневном введении животным ренина отмечается выраженный вазопрессорный эффект, в дальнейшем, несмотря на такие же дозы ренина, эффект бывает менее выражен, а затем животное перестает реагировать на него. Этот феномен, по мнению Goldblatt (1947), объясняется возникновением в крови животных вещества, нейтрализующего ренин, — антиренина. Это было подтверждено исследованиями Н. Т. Ковалевой (1953), когда после введения животному в кровь ренина, предварительно смешанного с сывороткой крови животного, переставшего на него реагировать, гипертензивный эффект не наблюдался.
Таким образом было установлено, что в юкстагломерулярном комплексе почки в условиях ее ишемии вырабатывается ренин, при дальнейшем его взаимодействии с белковыми компонентами плазмы образуется ангиотонин, вызывающий повышение артериального давления. Иначе говоря, почка оказывает воздействие на тонус артериальных сосудов, при изменении почечного кровообращения это воздействие проявляется артериальной гипертонией.
Однако дальнейшие исследования показали, что не только в условиях ишемии почки возникает гипертензивный эффект. При двусторонней нефрэктомии у животных, жизнь которых поддерживалась при помощи гемодиализа или перитонеального диализа, также возникает стойкая артериальная гипертония. В этих случаях не приходится говорить о выработке ренина, так как почечная паренхима отсутствует. На основании этих наблюдений возникла гипотеза о том, что почка наряду с ренином вырабатывает еще и гипотензивную субстанцию, отсутствие которой после двусторонней нефрэктомии вызывает артериальную гипертонию, вследствие невозможности нейтрализовать экстраренальные вазопрессорные вещества (норадреналин, дезоксикортикостерон и т. д.) (Grollman, 1953). Позднее Grollman удалось получить экстракты из почки, какие затем были выделены в чистом виде (кристаллическом). Уместно отметить, что эта так называемая гипотензивная субстанция оказывала гипотензивный эффект у больных гипертонией без каких-либо побочных реакций, какие обычно наблюдаются при терапии гипотензивными лекарственными препаратами.
Имеется мнение, что гипотензивная (антипрессорная) субстанция вырабатывается мозговым веществом почки. Видимо, при ишемии почки, когда возникает гипертония, наступает угнетение процессов, при которых вырабатывается антипрессорная субстанция, но вместе с тем усиливается продукция ренина. Эти два момента, по всей вероятности, и вызывают артериальную гипертонию. Несмотря на то что была получена так называемая антипрессорная субстанция, еще не ясен вопрос, обладает ли она сама гипотензивным эффектом или же она только нейтрализует прессорные вещества?
Как показали исследования Grollman (1953), Harrison (1955), большое влияние на почечную гипертонию оказывает нарушение электролитного баланса плазмы. Ограничение натрия в пище вызывает снижение артериального давления при почечной гипертонии, у здоровых же животных уменьшение количества натрия в пище не влияет на артериальное давление. Подобные же наблюдения были получены у животных после двусторонней нефрэктомии; повышение натрия в пищевом рационе ведет к повышению артериального давления. При недостатке калия возникает падение артериального давления как у здоровых, так и у животных с гипертонией. Нормализация артериального давления в этих случаях возникает лишь после введения кортизона. Все это еще раз подтверждает роль эндокринной системы в развитии почечной гипертонии.
В настоящее время установлено, что наряду с ренином и образуемым при его участии ангиотонином (ангиотензином) имеется ряд веществ: гидро-окситирамин, пиперидин, ферантразин и др. (Hamburger с сотр., 1965), которые обладают прессорным эффектом. Хотя эти вещества и не образуются в почке, однако нельзя исключить, что они играют роль в поддержании высоких цифр артериального давления у больных с почечной гипертонией.
Литература
Асратян Э. А. Изв. Научн. ин-та им. П. Ф. Лесгафта, 1934, 17—18, 221.
Великанова Л. К. Тканевые рецепторы осморегулиругощего рефлекса. Дисс.
Новосибирск, 1955.
ВихертА. М. и СеребровскаяЮ. А. Кардиология, 1962, 4, 10.
Гинецинский А. Г. В кн.: Тезисы докладов VIII съезда физиологов. М., 1955, стр. 170.
Гинецинский А. Г. В кн.: Доклады XX Международного конгресса физиологов.
М., 1956, стр. 235.
Гинецинский А. Г. Физиол. журн. СССР, 1959, 45, 761. Гинецинский А. Г. Физиологические механизмы водно-солевого равновесия. М.—Л., 1964.
ГинецинскийА. Г., БройтманА. Я. и Иванова Л. Н. Бюлл. эксп. биол
и мед., 1954, 38, 8.
Зарецкий И. И. Клиническая физиология и методы функциональной диагностики
почек. М., 1963.
Ковалева Н. Т. Арх. патологии, 1953, XV, 38.
Николаев П. Н. (1932—1934). Цит. по И. И. Зарецкому, 1963.
П ы т е л ь А. Я. Врач, дело, 1953, 3, 261.
ПытельА.Я. Лоханочно-почечные рефлюксы и их клиническое значение. М., 1959. Ш в а л е в В. Н. Иннервация почек. Л., АН СССР, 1965.
AstrupT. a. SterndorffJ. Proc. Soc. exp. Biol. Med., 1952, 81, 3, 675.
BargmannW. Histologie tmd mikroskopische Anatomie des Menschen. Stuttgart, 1956. В a r k e r E. S., S i n g e r R. В., E 1 k i n t о n J. R.a.CIarkJ. K. J. Clin. Invest., 1957,
36, 515.
В а г 11 e г F. C, M i 11 s I. H. a. G a n n D. S. J. Clin. Invest,, 1959, 38, 986. BoulangerP. etSiserte G. Acides amines libres et combines de l'urine humaine.
In Exposes annels de Biochimie medicale. Paris, 1956.
Braun-MenendezE. a. Page I. H. Science (New York), 1958, 127, 242. Braun-Menendez E., Fasciolo J. C, Leloir L. F., M u ft о z J. M. a. T a -
quini A. C. Renal Hypertension. Springfield, 1946. Chinard F. P. Renal function. Transactions of the third conference. Edit, by S. E.
Bradley. New York, 1952.
С u s h ny A. R. Secretion of urine. London, 1917. С u s h n у A. R. The secretion of urine. London, Edit. 2, 1926. D u n i h u e F. Anat. rec, 1949, 103, 442.
Elkinton J. R., S q u i r e s R. D. a. SingerR.B.J. Clin. Invest., 1951, 30, 381. Ellinger Ph. u. Hirt A. Zur Funktion der Nierennerven. Arch. exp. Path, und
Pharmakol., 1925, 106, 135. FryszmanA. Ztschr. Urol., 1927, 21, 481. F u с h s F. J. Urol., 1934, 32, 115.
GilmanA. a. BrazeauP. Am. J. Med., 1953, 15, 765. Goldblatt H. Physiol. Rev., 1947, 27, 120. Goldblatt H.,Hanzal R. F., Lynch J. a. Summerville W. W., J. exp.
Med., 1934, 59, 347.
GoormaghtighN. J. Pathol. a. Bact., 1945, 57, 392. GottschalkC. a. MylleM. Am. J. Physiol., 196, 927. GovaertsP. et Lambert P. P. J. Urol. med. chir., 1953, 59, 693. G г о 11 m a n A. a. Shapiro A.P.J. Clin. Invest., 1953, 32, 312. Hamburger J., Richet G., GrosnierJ. et Funck-Brentano J. L. L'in-
suffisance renale. Encyclopedie d'Urologie. T. IV. Berlin, 1962. Русский перевод.
Изд. Медицина, М., 1965.
Harrison Н. Е. а. Н а г г i s о n H. С. L. Clin. Invest., 1955, 34,1662. HartroftP. M. a. HartroftW. S. Anat. Rec, 1952, 112, 39. Henderson (1905). Цит. по И. И. Зарецкому, 1963. Jacobson L. O., Goldwasser E., Fried W. a. Plazak L. F. Trans. Ass. Am.
Physiol., 1957., 70, 305.
L a n di s E. Capillary pressure and capillary permeabilitv. Physiol. Rev., 1934, 14, 404. L a n d i s E. M. Am. J. med. Sci., 1937, 193, 297. Lassen a. Husfeldt (1934). Цит. по И. И. Зарецкому, 1963. L a 1 i e n О. M. a. P h i 11 i p s H. J. Urol., 1966, 95, 1, 90. Magnus (1900). Цит. по И. И. Зарецкому, 1963. MangazziniE . Quaderni di Urologia, 1962, 12, 2, 111. NashT.P. а. В e n e d i с t S. R. J. Biol. Chem., 1921, 48, 463. PappenheimerJ. Physiol. Rev., 1953, 33, 387.
Peter K. Untersuchungen tiber Bau und Entwicklung der Niere. Jena, 1909. Petersen a. Jaffe (1924). Цит. по И. И. Зарецкому, 1963. Pitts R. F. Federation Proc, 1948, 7, 418. Pitts R. F. Am. J. Med., 1958, 24, 745. Pitts R. F., L о t s p e i с h W. D., S с h i e s s W. А. а. А у e r J. L. J. Clin. Invest.,
1948, 27, 48. Polonovski C. et Colin J. Insuffisances congenitales du tubule renal chez Penfant.
T. I, Paris, 1957.
R e 1 m a n A. S. Am. J. Med., 1954, 17, 435.
R о b e r t s K. E., R a n d a 11 H. T. a. S a n d e r s H. J. Clin. Invest., 1954, 33, 960. R о 1 n i с k H. С a. S i n g e r P. L. J. Urol., 1947, 57, 834. R u у t e r J. H. С Ztschr. Zellforsch., 1925, 2, 242. S a h 1 i W. Pfluges Arch. Physiol., 1885, 36, 209. S с h w a r t z W. В., J e n s о n R. L. a. R e 1 m a n A. S. J. Clin. Invest., 1955, 34, 673.
Smith H. W. Principles of renal physiology. Oxford, 1956.
TigerstedtR. u. Bergman P. G. Skand. Arch. Physiol., 1898, 8, 223.
T о b i a n L. Physiol. Rev., 1960, 40, 280.
Tobian L. Circulation, 1962, 25, 189.
Trueta J., Barclay A. C, Daniel P.M., Franklin K. J. a. P r i с h a r d M. L.
Studies of the renal circulation. Oxford, 1947.
Ullrich K., Kramer K. a. BoylanJ. Progress in cardiovasc. diseases, 1961, 3, 394. Van der A. J., Mai vi n R. L., Wilde W. S. a. Sullivan L. P. Am. J. Physiol.,
1958, 195, 558.
V a n S 1 у к e D. D., P h i 11 i p s R. A., Hamilton B. P., Archibald R. M.,
D о 1 e V. P. a. EmersonK. Trans. Ass. Am. Physiol., 1944, 58, 119.
V о g e 1 G. u. H e у m E. Klin. Wschr., 1955, 33, 627. WachsteinM. XX Congress Internat. Physiol., 1956, 938. W i n t о n F. R. J. Physiol., 1931, 72, 49.
Wirz H. Helvetica Physiol. Acta, 1952, 11, 20.
Wirz H. Helvetica Physiol. Acta, 1956, 14, 353.
Глава четвертая
Физиология
верхних мочевых путей
и мочевого пузыря
Физиология верхних мочевых путей
Функция почки тесно связана с функцией верхних мочевых путей. При нарушении последней очень быстро страдает функциональная способность почки. Достаточно сказать, что большое число почечных заболеваний зависит от нарушения функции почечных чашечек, лоханки и мочеточника.
Верхние мочевые пути обеспечивают эвакуаторную функцию, т. е. транспортируют мочу, вырабатываемую почкой, в мочевой пузырь. Функция верхних мочевых путей тесным образом связана с функцией почки и функцией мочевого пузыря и обеспечивается нервно-мышечным аппаратом почечных чашечек, лоханки и мочеточника. Нервная система верхних мочевых путей взаимосвязана с нервами почки, ее сосудами и с нервами мочевого пузыря. Этим и объясняется их динамическое взаимодействие (Ю. А. Пытель, 1960). Мышечная система верхних мочевых путей, условно делимая на мышцы почечных чашечек, лоханки и мочеточника, представляет собой на самом деле единую стройную мышечную архитектонику, построенную по принципу взаимно переплетающихся спиралей. Этот тип строения мышечной оболочки верхних мочевых путей обеспечивается за счет переплетения мышечных пучков, идущих в различных направлениях и переходящих из одного слоя в другой (Schneider, 1938; А. Я. Пытель, 1950; С. А. Баялова, 1951).
Для удобства описания физиологии верхних мочевых путей мы подразделяем этот единый динамический процесс на функцию почечных чашечек, лоханки и мочеточника.
Основным процессом, обеспечивающим транспорт мочи из выводных канальцев почки, является функция почечных чашечек. Почечные чашечки в отличие от других отделов верхних мочевых путей имеют определенные сфинктерные мышечные группы, которые представляют собой циркулярное строение единой мышечной системы почечных чашечек, сосочка и шейки. Мышечные группы одного сфинктера переходят в другой, спирально охватывая чашечку.
Поступление мочи из почечной паренхимы в чашечку, а затем в почечную лоханку осуществляется в два этапа: фазой диастолы—накопления мочи в чашечке и фазой систолы — опорожнения чашечки в лоханку (рис. 28). Во время диастолы m. levator fornicis находится в расслабленном состоянии и сосочек как бы приближен к чашечке за счет укорочения его свода. М. sphincter fornicis Henle расслаблен — раскрыт, чему способствует сокращение верхней части m. longitudinalis. Одновременно с этим m. sphincter calycis Disse находится в сокращенном состоянии — замкнут, что предупреждает проникновение мочи в чашечку из лоханки. В дальнейшем происходит еще большее расслабление m. sphincter fornicis и некоторое сокращение m. levator fornicis, следствием чего является увеличение объема почечной чашечки при еще закрытом, сокращенном m. sphincter calycis. Это способствует созданию вакуума под сосочком и в результате этого моча из собирательных канальцев поступает в чашечку. Наличие относительно отрицательного давления в чашечке в момент диастолы необходимо, так как имеющееся гидравлическое давление в выводных канальцах (максимальное до 20—25 мм рт. ст.) недостаточно для транспорта мочи в мочевые пути, так как в лоханке оно при нормальных условиях достигает 50 мм рт. ст.
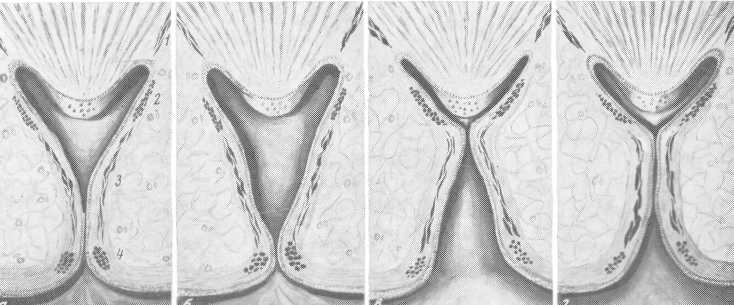
Рис. 28. Уродинамика почечных чашечек.
а, б — диастоличеокая фаза; в, г — систолическая фаза.
При отсутствии отсасывающего эффекта в чашечке был бы невозможен транспорт мочи из почки. После наполнения почечной чашечки мочой наступает фаза систолы — опорожнения. При этом m. sphincter calycis расслабляется — шейка чашечки раскрывается, одновременно с этим замыкается m. sphincter fornicis, чему содействует расслабление верхней части m. longi-tudinalis и еще большее сокращение т. levator fornicis. В итоге стенки чашечки прижимаются к сосочку, что предотвращает проникновение мочи в выводные канальцы. В это же время нижняя часть m. longitudinalis сокращается, способствуя опорожнению чашечки и еще большему расслаблению m. sphincter calycis и тормозя тем самым фазу накопления. По мнению Na-rath (1951), процессу опорожнения чашечки способствует содействие мышечных пучков, отходящих от m. sphincter calycis в сторону лоханки (m. recludor calycis) (А. Я. Пытель, 1950, 1959).
Периодическое чередование фаз систолы и диастолы обеспечивает транспорт мочи из почки в почечную лоханку. Следует указать, что деятельность почечных чашечек зависит от давления в почечной лоханке и самих чашечек, что регулируется барорецепторами. Скорость и сила динамики почечных чашечек зависят от диуреза и соответственно от величины давления. Опорожнение почечной чашечки наступает лишь в том случае, когда давление мочи в ней станет пороговым, обусловливающим рефлекс на ее опорожнение. При повышении давления в почечной лоханке выше нормально допустимого шейка почечной чашечки замкнута за счет сокращения т. sphincter calycis, что предупреждает проникновение мочи в чашечку и возникновение пиелоренального рефлюкса. Однако при все продолжающемся повышении внутрилоханочного давления происходит перераздражение барорецепторов. При этом раскрывается m. sphincter calycis и моча из лоханки устремляется в чашечку. Давление в почечной лоханке снижается и одновременно с этим включается форникальный аппарат, обеспечивающий резорбцию мочи, чем еще больше снижается внутрилоханочное давление. Поскольку объем почечных чашечек относительно большой, то при проникновении мочи из лоханки давление в них не будет предельным, что препятствует возникновению лоханочно-почечных рефлюксов. Последние могут возникнуть лишь в том случае, когда после проникновения мочи из лоханки в чашечки давление в лоханочно-чашечной системе будет увеличиваться и станет предельным. Однако возникновение лоханочно-почечных рефлюксов возможно и при невысоких цифрах внутрилоханочного давления, при дискинезии, когда из-за несоразмеренных сокращений мышц почечных чашечек может возникнуть в них предельно высокое давление.
Несмотря на тесную взаимосвязь функции почечных чашечек и лоханки, не выявлена определенная синхронность в деятельности малых почечных чашечек. Сокращение различных чашечек по скорости и интенсивности, как правило, различно, что, видимо, связано с процессами, происходящими в различных отделах почечной паренхимы. Такое неравномерное функционирование почечных чашечек было установлено в последнее время при помощи урорентгенокинематографии, которая позволила выяснить ряд физиологических закономерностей верхних мочевых путей (Bodner, Howard, Kaplan, 1958).
Suyama (1961) установил, что имеется определенная закономерность в функции верхней и средней чашечек, т. е. начало их сокращений совпадает, тогда как сокращение нижней чашечки не является синхронным. Уместно отметить, что динамика опорожнения почечных чашечек у различных людей различна и зависит от ряда причин, среди которых в первую очередь следует выделить величину диуреза, качество и количество пищи (что, видимо, связано с изменением функции почки в зависимости от алиментарного фактора) и особенно положение тела. В вертикальном положении тела сокращение чашечек становится чаще и короче. В процессе динамики опорожнения верхних мочевых путей некоторые чашечки длительное время остаются спастически сокращенными, другие же, наоборот, наполнены мочой. При полиурии, при сокращении почечной лоханки иногда удается наблюдать с помощью урорентгенокинематографии обратное проникновение мочи в чашечки, что зависит от повышенного внутрилоханочного давления, о чем говорилось выше. Hanley (1956, 1957), Fey, Truchot, Noix (1958) установили, что диастолическая фаза почечной чашечки в среднем продолжается 4 секунды, а систолическая — 3 секунды. Между систолой и диастолой имеется пауза, которая длится 5—10 секунд. Исследования Mitsuya, Asai, Suyama, Sai, Hosoe (1962) показали, что движения форникса и шейки чашечки согласованы. Время сокращения в зоне форникса колеблется в пределах P/g—25/8, расслабления — 1—22/8, а в зоне шейки чашечки сокращения— 1—25/8, расслабления —15/8—З1/^ секунды. Эти же авторы установили, что наиболее распространенным у здоровых людей тип деятельности почечной чашечки следующий: покой, систола, диастола, покой. Более редко может наблюдаться другой распорядок динамических фаз в чашечках: покой, сокращение, расширение, редукция, покой, или: покой, сокращение, редукция, покой, или: покой, расширение, сокращение, расширение, редукция, покой.
Функция почечных чашечек тесным образом связана с функцией почечной лоханки, что и заставило объединить эти образования в одну динамическую систему — лоханочно-чашечная система. Лоханка аналогична
чашечкам, функционирует по принципу детрузор — сфинктер. Роль сфинктера в лоханке выполняет лоханочно-мочеточниковый сегмент.
Для того чтобы лучше уяснить вопросы физиологии почечной лоханки и мочеточника, уместно остановиться на теории Фукса о цистоидном строении верхних мочевых путей. Fuchs (1934) рассматривает все верхние мочевые пути как отдельные детрузорно-сфинктерные образования: чашечки, где роль детрузора выполняет верхняя группа мышц (rn. sphincter fornicis, m. levator fornicis и верхняя часть m. longitudinalis), а роль сфинктера — m. sphincter calycis и нижняя часть m. longitudinalis. Чашечки Fuchs именует эпицистоидами. Следующей детрузорно-сфинктерной секцией является лоханка с лоханочно-мочеточниковым сегментом, где последний играет роль сфинктера. В мочеточнике имеется от двух до четырех цистоидов, которые обеспечивают транспорт мочи из почечной лоханки в мочевой пузырь. В лоханочно-мочеточниковом сегменте и в мочеточнике нет мышечных сфинктеров, что подтверждено многочисленными морфологическими исследованиями (Fuchs, 1931; Schneider, 1938; Renyi-Vamos, Balogh, Szendroi, 1948; Narath, 1951; А. Я. Пытель, 1950; Begg, 1946; С. А. Баялова, 1952; Ю. А. Пы-тель, 1960), однако эти участки мочеточника функционируют по принципу сфинктеров. Удалось установить, что в местах так называемых физиологических сфинктеров располагаются кавернозноподобные образования, выполняющие роль гидравлических жомов и обеспечивающие вместе с сегментарным сокращением мускулатуры этой зоны мочеточника функцию сфинктера, подобно тому, как это имеет место в uvulae vesicae (Ю. А. Пытель, 1960). Подробнее о строении и функции кавернозноподобных образований будет изложено в разделе о функции мочеточника. Теория Фукса в настоящее время подтверждена большим числом клинико-рентгенологи-ческих наблюдений. Особенно большую пользу в этом отношении сыграла урорентгенокинематография, которая позволила достоверно установить, что динамика опорожнения верхних мочевых путей осуществляется по принципу цистоидов (Fey с сотр., 1958).
В лоханке, как и в чашечке, различают фазы систолы и диастолы. Систола продолжается 2—-3 секунды, диастола — 10—15 секунд. Систоле предшествуют мелкие волнообразные сокращения лоханки (пресистолическая фаза диастолы), вслед за которыми наступает резкое ее сокращение. Наполнение лоханки мочой происходит постепенно, за счет сокращения почечных чашечек. Емкость лоханки превышает емкость чашечек, в связи с чем чашечки сокращаются чаще, чем лоханка. Fey с сотр. (1964) наблюдал на 26 систол чашечек только 9 сокращений лоханки. Сокращения чашечек не продолжаются на лоханку и мочеточник, они автономны. Нельзя представлять себе лоханку исключительно как вместилище для мочи, поступающей из чашечек. В момент диастолы происходит активное растяжение мышц лоханки, обусловливающее в ней отрицательное давление, что наряду с сокращением чашечек облегчает поступление мочи в лоханку. Лоханка никогда во время систолы не опорожняется полностью. В момент ее сокращения моча устремляется в верхнюю часть мочеточника (цистоид-I), так как в это время открыт лоханочно-мочеточниковый сегмент, а также частично в сторону чашечек, шейки которых находятся в раскрытом состоянии. Однако зона сосочка и свода закрыта за счет сокращения верхней группы мышц чашечки, что предупреждает развитие лоханочно-почечного рефлюк-са. Такого рода динамику лоханочно-чашечной системы наблюдал Fey с сотр. (1964) при урорентгенокинематографии. То положение, что лоханка в момент систолы никогда не опорожняется полностью, можно объяснить следующим образом. Динамическая деятельность верхних мочевых путей и, в частности, лоханки построена на основе барорецепции. Фаза систолы возникает в том случае, когда давление в лоханке стало пороговым, а прекращение сокращения характеризуется достижением давления в верхней части мочеточника (цистоид-I) до пороговых цифр с одновременным падением давления в лоханке. В результате этого возникает закрытие лоханочно-мочеточникового сегмента и прекращение систолы лоханки еще задолго до ее полного опорожнения. Это связано с тем, что емкость верхней части мочеточника (цистоид-I) значительно меньше емкости лоханки и рефлекс возникает на закрытие лоханочно-мочеточникового сегмента после того, как наполнится мочой эта часть мочеточника, что по времени совпадает с прекращением систолы лоханки. От емкости верхней части мочеточника (цистоид-I) зависит тонус лоханки. При внутрипочечном типе строения лоханки тонус лоханочно-чашечной системы бывает более высоким, так как при каждом ее сокращении почти вся моча будет поступать в мочеточник. В то время как при ампулярном типе почечной лоханки (внепочечном) тонус ее будет ниже опять-таки за счет того, что емкость верхней части мочеточника (цистоид-I) намного меньше, чем емкость лоханки такого типа. Поэтому внепочечная лоханка при ее сокращении опорожняется намного меньше, чем лоханка малой емкости внутрипочечного типа. Более низкий тонус лоханки создает условия, не позволяющие переполнять и перерастягивать верхнюю часть мочеточника (цистоид-I). Таким образом, тонус лоханки зависит от емкости верхней части мочеточника (цистоид-I) в первую очередь и от формы и объема самой лоханки, а также от диуреза. Низкий тонус лоханки внепочечного типа компенсируется высоким тонусом мочеточника, сокращение которого происходит чаще и с большей силой, нежели мочеточника при лоханке малой емкости внутрипочечного типа. В зависимости от типа почечной лоханки, ее тонуса будет и различным пороговое внутрилоханочное давление, регистрация которой барорецепторами обусловливает начало систолы лоханки. При внутрипочечном типе оно будет ниже, чем при внепочечном. Наблюдаемое проникновение мочи в чашечки при сокращении лоханки надо рассматривать как своеобразную защитную реакцию, какая предотвращает перенаполнение верхней трети мочеточника. Видимо, проникновение мочи в чашечки в момент систолы лоханки зависит от тонуса последней. Если он высокий и сокращение лоханки происходит в короткий промежуток времени, то в этом случае чашечки примут большую часть мочи, чем снизится внутрилоханочное давление и соответственно с этим не возникнет резкого перенаполнения верхней части мочеточника. Если же тонус лоханки будет на данном этапе низким, то степень проникновения мочи в чашечки будет меньшей, чем это имело место при высоком тонусе почечной лоханки. Иначе говоря, наполнение чашечек мочой при сокращении лоханки есть рефлекторный процесс, зависящий от тонуса лоханки, определяющего в известной мере внутрилоханочное давление и соответственно скорость динамики опорожнения мочи по верхним мочевым путям.
Говоря о функции почечной лоханки, мы не освещали особенностей анатомического строения мочеточника и соответственно его функции, хотя эти два органа в этом отношении теснейшим образом связаны. В мочеточнике нет мышечных сфинктеров, однако места физиологических сужений функционируют по принципу сфинктеров, вследствие этого указанные сфинктероподобные перетяжки стали условно именовать „физиологическими" сфинктерами. Исследования последних лет позволили установить, что в местах так называемых физиологических сфинктеров располагаются своеобразные сосудистые образования. Они располагаются в зоне лоханочно-мочеточникового сегмента, на границе верхней и средней трети, средней и нижней трети и в интрамуральном пузырном отделе мочеточника (Ю. А. Пытель, 1960).
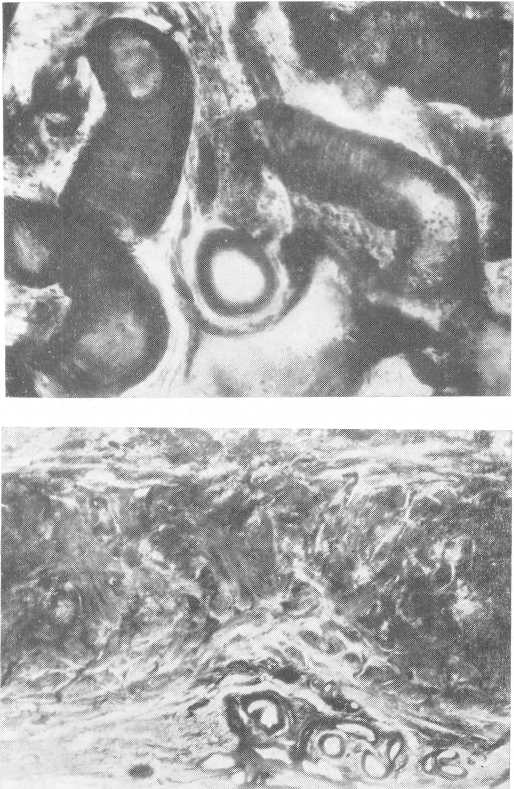
Рис. 29. Кавернознопо-добное образование в лоханочно-мочеточниковом сегменте. Микрофото.
Рис. 30. Кавернознопо-добное образование на границе мышечной и слизистой оболочек средней части мочеточника.
Эти образования представляют собой скопление в подслизистой и межмышечной соединительной ткани мочеточника большого числа сосудов, весьма крупных по своему калибру (рис. 29, 30). Эти крупные сосуды располагаются близко друг к другу, т. е. почти рядомг имеют извитый ход и напоминают строение пещеристых тел. То, что эти сосудр1стые образования, именуемые кавернозноподобными, располагаются в зоне физиологических сфинктеров и наряду с ними имеются обычные сосуды в таком же количестве, как и в других участках мочеточника, заставило высказать следующую мысль: вряд ли они выполняют одну лишь роль васкуляризации, тем более, что очень большой их размер и их обилие не соответствуют условиям питания весьма небольших участков мочеточника. Было высказано мнение, что кавернозноподобные образования выполняют роль гидравлических жомов в момент сокращения этих зон мочеточника. При наполнении кровью кавернозноподобного образования резко увеличивается давление на стенку мочеточника, что при определенном пороге раздражения вызывает сегментарное спастическое сокращение мускулатуры соответствующего участка мочеточника; это вместе с так называемой гидравлической муфтой кавернозноподобного образования обеспечивает полное замыкание просвета мочеточника. Опорожнение кавернозноподобного образования ведет к снижению давления на стенку мочеточника, расслаблению мускулатуры зоны физиологического сфинктера и тем самым к открытию просвета мочеточника.
Таким образом, так называемые физиологические сфинктеры мочеточника обеспечиваются двумя компонентами: гидравлическим жомом кавер-нозноподобного образования и спастическим сокращением гладкой мускулатуры этой зоны. При цистоидном строении верхних мочевых путей нет перистальтических сокращений, а имеют место периодически сменяющие друг друга заполнения и опорожнения последовательно расположенных его участков. В каждом мочеточнике имеется несколько динамических цистоидных секций, чаще три, реже две или четыре. Каждой такой секции или цистоиду присущи детрузорно-сфинктерные закономерности, столь хорошо выраженные и изученные в отношении мочевого пузыря. Динамические секции —- цистоиды мочеточника — функционируют таким образом, что в то время как одна из них находится в сокращенном состоянии, другая, расположенная рядом с ней, — в расслабленном, расширенном состоянии. Корреляция тонуса между сфинктером и детрузором гарантирует опорожнение каждого цистоида мочевых путей. В мочеточнике ритмическая деятельность цистоидов чередуется; обычно в то время, когда, например, верхний цистоид находится в фазе наполнения, средний находится в фазе сокращения, а нижний — вновь в фазе наполнения — расширения и наоборот. Цистоиды разделены зонами, где располагаются кавернозноподобные образования, осуществляющие роль сфинктеров. Деятельность этих образований может быть представлена следующим образом (рис. 31). После поступления мочи из почечной лоханки в верхнюю треть мочеточника (цистоид-I) наступает фаза заполнения сосудов кавернозноподобного образования кровью и закрытие просвета мочеточника в лоханочно-мочеточниковом сегменте. После этого начинается заполнение мочой почечной лоханки и одновременно с этим за счет сокращения мускулатуры верхней трети мочеточника (цистоид-I) продвижение мочи в среднюю треть мочеточника (цистоид-И), чему предшествует опорожнение кавернозноподобного образования на границе верхней и средней трети и раскрытие просвета мочеточника этой зоны. Иначе говоря, функция кавернозноподобных образований так называемых физиологических сфинктеров в расположенных рядом цистоидах антагонистична. Если кавернозноподобное образование вышележащего цистоида заполнено кровью и просвет мочеточника закрыт, то кавернозноподобное образование нижележащего цистоида опорожнилось от крови и просвет мочеточника в этой зоне зияет. В следующем цистоиде повторяется процесс, аналогичный первому, т. е. кавернозноподобное образование заполнено кровью и вследствие этого просвет мочеточника закрыт. Функция так называемых физиологических сфинктеров во многом зависит от барорецепции, как и функция всех отделов верхних мочевых путей.
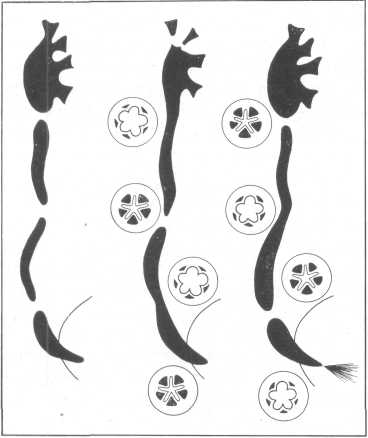
Рис. 31. Схема цистоидиых секций верхних мочевых путей.
По мере заполнения почечной лоханки мочой увеличивается давление на ее стенки и по достижении соответствующего порога раздражения возникают импульсы от барорецепторов. В ответ на это возникает опорожнение кавернозноподобного образования, раскрытие просвета мочеточника в зоне лоханочно-мочеточникового сегмента, сокращение мускулатуры почечной лоханки и в итоге осуществляется продвижение мочи в верхнюю часть мочеточника—в цистоид-I. После того как лоханка опорожнилась частично от мочи, давление в ней снизилось и наступила фаза покоя или диастолы, возникает рефлекторное заполнение сосудов кавернозноподобного образования лоханочно-мочеточникового сегмента, что приводит к закрытию просвета мочеточника в этой зоне. Этот рефлекс возникает не со стороны лоханки, а со стороны верхней трети мочеточника — цистоида-I. Прекращение систолы лоханки происходит рефлекторно, как только давление мочи в верхней части мочеточника — в цистоиде-1 — станет пороговым и в ответ на это произойдет заполнение кровью сосудов кавернозноподобного образования лоханочно-мочеточникового сегмента и закрытие просвета мочеточника. В это же время наступает фаза систолы в нрхжележащем цистоиде-1, где давление мочи на стенку мочеточника стало пороговым. Рефлекторно происходит опорожнение кавернозноподобного образования и вследствие этого раскрытие просвета мочеточника, продвижение мочи в цисто-ид-П. Таким образом, раскрытие так называемого физиологического сфинктера происходит в результате рефлекса с барорецепторов вышележащего цистоида на кавернозноподобное образование, расположенное ниже. Закрытие же просвета мочеточника осуществляется также рефлекторно, но уже с барорецепторов нижележащего цистоида на кавернозноподобное образование, расположенное выше него.
В нижнем цистоиде, видимо, рефлекс на заполнение сосудов кавернозноподобного образования и закрытия просвета мочеточника в области устья возникает по мере падения давления в этом отделе. Можно, конечно, предположить, что рефлекс на закрытие просвета мочеточника в области «физиологических» сфинктеров возникает и с выше- и с нижележащего цистоида во всех секциях мочеточника. С вышележащего — при падении давления, а с нижележащего — при увеличении давления. Однако более логично считать, что рефлекс на закрытие просвета мочеточника возникает с нижележащего цистоида, когда давление мочи на его стенку станет пороговым, тем более, что ввиду различного объема рядом расположенных цистоидов регистрация повышения и падения давления в них будет разновременной, а следовательно, и рефлексы на один и тот же процесс будут возникать в различные короткие промежутки времени, что безусловно повлечет за собой возникновение дискинезии. Рефлекторную реакцию с одного цистоида и на закрытие и на раскрытие просвета мочеточника можно допустить лишь в самом мощном нижнем цистоиде. Этой функциональной особенностью можно объяснить существование более мощной мышечной оболочки и наличия большого количества рецепторов и целых рецептор-ных полей (Ю. А. Пытель, 1955, 1960). Видимо, высокий тонус нижнего цистоида мочеточника защищает вышележащие отделы мочевых путей и особенно почку от нарушений в корреляционной деятельности мочевого пузыря и мочеточника, когда имеется вероятность возникновения пузырно-мочеточникового рефлюкса.
Роль барорецепции в динамике опорожнения мочи из верхних мочевых путей иллюстрируется наблюдениями над больными с нефростомой при производстве им антеградной пиелографии. Введение контрастной жидкости в малом количестве и под низким давлением не вызывает раскрытия лоханочно-мочеточникового сегмента, но достаточно повысить давление в лоханке на 40—50 мм рт. ст., как контрастное вещество начинает проникать в верхнюю треть мочеточника — в цистоид-I, а затем по мочеточнику, выявляя сегментарно фазы систолы и диастолы соответственно тому или другому цистоиду. Отдавая должное барорецепции, все же может возникнуть мысль, что на динамику почечной лоханки и мочеточника влияет непосредственно моча, т. е. рефлексы осуществляются с хеморецепторов. Ни в коей мере нельзя отрицать роль хеморецепторов в верхних мочевых путях, однако вряд ли они играют ведущую роль в рефлекторном процессе продвижения мочи по лоханке и мочеточнику, так как заполнение их контрастной жидкостью, физиологическим раствором, дистиллированной водой и т. д. при антеградной пиелографии не вызывает заметного нарушения динамики. Это и позволяет прийти к выводу, что рефлекторные процессы опорожнения мочи из верхних мочевых путей осуществляются в основном за счет барорецепции. Следует отметить, что степень сокращения различных цистоидов неодинакова; она находится в зависимости от тонуса мускулатуры отдельных участков мочеточника. Так, тонус верхнего и нижнего цистоидов приблизительно одинаковый, давление в этих участках мочеточника достигает 40—50 мм рт. ст., а в среднем цистоиде оно низкое и равно 20—25 мм рт. ст. Эти наблюдения подтверждаются и урорентгено-кинематографическими исследованиями. В верхнем и нижнем цистоиде происходит сильное и быстрое сокращение мочеточника, когда в среднем цистоиде систола бывает представлена в виде медленной вялой волны (Mitsura, Asai, Suyama, Hosoe, 1962). В области мочеточникового устья не имеется каких-либо мышечных сфинктеров, и закрытие его просвета также осуществляется при участии кавернозыоподобного образования и мышечной стенки мочевого пузыря, которая не замыкает просвет мочеточника, а лишь способствует поддержанию высокого тонуса его мускулатуры. Говоря о роли мочевого пузыря в динамике опорожнения верхних мочевых путей, следует указать на его активное участие в этом процессе (Lapides, 1948; Kiil, 1957; Tonagho и Pugh, 1963). Подобно почечной лоханке и цисто-идам мочеточника мочевой пузырь после своего опорожнения начинает активно расслабляться, создавая при этом относительное отрицательное давление, что способствует более легкому поступлению мочи из мочеточников.
Единство анатомического строения, обилие нервных взаимосвязей почки, лоханочно-чашечной системы, мочеточника и мочевого пузыря объясняет всю сложную и стройную функциональную систему верхних мочевых путей.
Физиология мочевого пузыря
Мочевой пузырь выполняет ряд функций, обеспечивающих нормальную эвакуацию мочи из организма. Одна из них — это акт мочеиспускания. Акт мочеиспускания обсуловлен сокращением детрузора в комплексе с сокращением мышц передней брюшной стенки и урогенитальной диафрагмы в сочетании с раскрытием так называемого замыкательного аппарата мочевого пузыря — пузырных сфинктеров. Представление о том, что замыка-тельный аппарат мочевого пузыря состоит из двух сфинктеров: внутреннего, содержащего гладкие мышцы, и наружного, содержащего поперечнополосатые, — схематично и глубоко условно.
Внутренний сфинктер мочевого пузыря представляет собой продолжение мышечных волокон детрузора, которые переходят resp., сливаются с продольной мускулатурой задней уретры. Эти мышечные волокна детрузора образуют утолщение в области заднего сегмента шейки пузыря и носят
название sphincter trigonalis. В момент сокращения этого сфинктера выход из мочевого пузыря замыкается вследствие прижатия задней утолщенной части шейки пузыря к передней стенке мочеиспускательного канала. В этой области нет циркулярных мышц, свойственных сфинктерам (Hutch, 1965).
Наряду с указанным механизмом в замыкательном аппарате мочевого пузыря принимают большое участие и другие анатомо-физиологические структуры. Так, из продольных наружных волокон задней стенки детру-зора образуется петля вокруг уретры, тотчас у ее отхождения от мочевого пузыря. Позади orificium urethrae internum располагается uvula vesicae — образование, представляющее собой венозное сплетение, находящееся между мышечными и соединительнотканными элементами этой области и имеющее вид валика.
При наполнении мочевого пузыря мочой рефлекторно происходит наполнение кровью венозного сплетения uvula vesicae, вследствие чего размеры его увеличиваются. Одновременно с этим в результате растяжения детрузора образованная из его волокон петля смещается книзу и прижимается к увеличенной uvula vesicae; это приводит к закрытию внутреннего отверстия уретры.
В динамике опорожнения мочевых путей большая роль принадлежит сосудистым образованиям, выполняющим роль так называемых физиологических сфинктеров. В мочеточнике — это кавернозноподобные образования, в мочевом пузыре — uvula vesicae.
Известную роль в замыкательном аппарате мочевого пузыря играет продольная мускулатура задней уретры. При сокращении этой мускулатуры задняя уретра укорачивается, тогда как просвет ее расширяется, и тем самым создаются условия для свободного выделения мочи из мочевого ттаыря. Функцию продольной мускулатуры уретры в акте мочеиспускания ь.ожно наблюдать на уретрограммах. На восходящей уретрограмме, когда замыкательный аппарат пузыря закрыт, задняя уретра представлена тонкой вытянутой трубкой, тогда как на нисходящих уретрограммах, выполненных в момент мочеиспускания, этот отдел мочеиспускательного канала оказывается расширенным и укороченным.
Одним из основных функционально-анатомических образований замы-кательного аппарата мочевого пузыря является наружный сфинктер, состоящий из поперечнополосатой мускулатуры. Этот сфинктер концентрически охватывает уретру от простатического до мембранозного ее отдела. Мышечные волокна наружного сфинктера переплетаются с мышцами промежности, которые принимают участие в акте мочеиспускания. Впереди к наружному сфинктеру примыкает m. bulbocavernosus, который в результате своего сокращения обеспечивает удаление последней порции мочи из уретры после завершения акта мочеиспускания.
Замыкательный аппарат мочевого пузыря у женщин устроен в принципе так же, как и у мужчин, с той лишь разницей, что мышечные элементы гладкой и поперечнополосатой мускулатуры расположены очень тесно друг к другу. Наружный сфинктер женской уретры охватывает проксимальный ее отдел циркулярно, располагаясь вокруг гладкой мускулатуры.
Помимо указанных запирательных механизмов мочевого пузыря, значительная роль в удержании мочи принадлежит фасциальным и связочным образованиям таза, фиксирующим уретру. Связочный аппарат таза и m. pubovesicalis фиксируют мочеиспускательный канал к лонному сочленению; это создает острый угол впадения уретры в мочевой пузырь при его наполнении мочой, что содействует удержанию мочи.
Роль фиксирующего аппарата уретры в акте мочеиспускания и удержании мочи демонстративно видна на урорентгенокинематографических фильмах. Перед мочеиспусканием происходит опущение дна пузыря, что связано с расслаблением урогенитальной диафрагмы и увеличением внут-рибрюшного давления. При мочеиспускании, особенно у женщин, происходит заметное изменение пузырно-уретрального угла, он из острого становится тупым. При прекращении мочеиспускания дно пузыря резко приподнимается, пузырно-уретральный угол становится острым, и это совпадает с разрывом струи мочи в уретре. В случае нарушения связочного аппарата уретры, особенно у женщин, изменяются взаимоотношения уретры и дна пузыря, что может быть причиной неудержания мочи.
Исключительно большая роль в функционировании мочевого пузыря принадлежит нервной системе. Иннервация пузыря обеспечивается из трех источников: из симпатической нервной системы — nn. hypogastrici; парасимпатической — nn. pelvici и спинного мозга — nn. pudendi. Nn. hypogastrici и nn. pelvici и образуют plexus vesicalis, которое иннервирует детрузор и внутренний сфинктер. Раздражение nn. hypogastrici вызывает сокращение сфинктера и расслабление детрузора, а раздражение nn. pelvici, наоборот, сокращение детрузора и расслабление сфинктера. Концепция Elliot (1907) об антагонистической иннервации мочевого пузыря в последнее время подвергается пересмотру. ТогЬеу(1963) считает, что основная роль в рганервации мочевого пузыря принадлежит парасимпатической нервной системе. Lapides, Hodgson, Boyd, Shook, Lichtwardt (1958) вообще отрицают участие симпатршуса в акте мочеиспускания. Saccomauso (1944) доказывает, что симпатический нерв осуществляет иннервацию только области мочепузырного треугольника. Иннервация наружного сфинктера осуществляется nn. pudendi.
Наряду с указанными иннервационными приборами мочевой пузырь обладает значительным интрамуральным ганглионарным аппаратом, который в основном располагается в области дна; он обеспечивает некоторый автоматизм мочевого пузыря. Это, например, подтверждается клиническими наблюдениями, когда у больных восстанавливается акт мочеиспускания после перерыва спинного мозга.
При увеличении объема мочевого пузыря и соответственно увеличении внутрипузырного давления возникают афферентные импульсы в центральную нервную систему, исходящие с механо- и барорецепторов. Начальным сигналом, вызывающим раздражение рецепторов мочевого пузыря, является растяжение его стенок, изменение тонуса детрузора. Ведущая роль в регистрации состояния тонуса детрузора принадлежит механорецепто-рам (В. М. Хаютин, 1961). После поступления первых афферентных импульсов в центральную нервную систему возникает усиление тонуса наружного сфинктера и одновременно с этим торможение детрузора, что ведет к снижению его тонуса и способствует дальнейшему поступлению мочи в пузырь из верхних мочевых путей. Возможное поступление нескольких капель мочи в заднюю уретру в этот момент рефлекторно вызывает еще большее сокращение наружного сфинктера (В. М. Хаютин). Однако это последнее мало вероятно при появлении первых позывов к мочеиспусканию; в пользу этого говорят данные цистографии и урорентгенокинема-тографии, когда не наблюдается проникновения контрастной жидкости в заднюю уретру при позыве к мочеиспусканию.
При достижении нормальной емкости мочевого пузыря, равной 200—-300 мл, поступает усиленный поток афферентных импульсов с механо- и барорецепторов в спинальные центры мочеиспускания (Ьц—Ьш и Sn—Srv). Соответственно возвращающиеся из боковых рогов эфферентные импульсы к детрузору вызывают его сокращение, в результате чего резко возрастает внутрипузырное давление, достигающее 60 см вод. ст. Сокращение детрузора вызывает укорочение и раскрытие шейки мочевого пузыря, чему способствует опорожнение от крови uvula vesicalis и расширение и укорочение задней уретры. Одновременно с этим из передних рогов спинальных центров через nn. pudendi поступают импульсы, расслабляющие наружный сфинктер.
Начало акта мочеиспускания сопровождается снижением внутрипузырного давления за счет поступления части мочи в уретру. Соответственно с этим уменьшается раздражение рецепторов и может прекратиться мочеиспускание. Однако этого не происходит, так как поступающая моча в заднюю уретру служит автоматическим возбудителем, подкрепляя и усиливая рефлекторное сокращение пузыря. Само же сокращение детрузора раздражает механорецепторы, чем еще больше подкрепляет рефлекс на опорожнение пузыря и расслабление сфинктеров (В. М. Хаютин). По окончании акта мочеиспускания замыкается наружный сфинктер, тогда как расслабление детрузора ведет к закрытию шейки пузыря.
Благодаря связям спинальных центров с головным мозгом — корой, гипоталамусом, средним мозгом, варолиевым мостом — функция высших отделов центральной нервной системы заключается в контроле порогов возбуждения рефлексов мочеиспускания. Усилием воли человек может вызвать акт мочеиспускания при малом количестве мочи в мочевом пузыре, а также подавить ощущение позыва к мочеиспусканию и даже прервать последнее.
Наряду с тем, что мочевой пузырь является резервуаром для мочи и одним из основных органов, участвующих в акте мочеиспускания, ему присущи и другие, не менее важные функции. Одной из них является функция регулирования и обеспечения нормального пассажа мочи иг верхних мочевых путей. Поступление мочи из мочеточников в мочевой пузырь — сложный рефлекторный процесс. Он происходит не только за счет деятельности мускулатуры мочеточников; большая роль в этом принадлежит функции мочевого пузыря. Нарушение функции мочевого пузыря довольно быстро приводит к расстройству функции верхних мочевых путей и почек, что способствует развитию в них болезненных процессов. Повседневные наблюдения за больными с нарушенной функцией мочевого пузыря подтверждают это.
Мочевой пузырь — это не пассивный резервуар, который растягивается по мере поступления в него мочи. Тесные нервные связи между мочевым пузырем и верхними мочевыми путями обеспечивают рефлекторную деятельность всех органов мочевыделительной системы. Именно рефлекторная регуляция функции мочевыводящих путей создает наиболее оптимальные условия для нормального пассажа мочи (Longworthy, Kolb и Lewis, 1940).
Нормальный пассаж мочи из верхних мочевых путей обеспечивается не только силой мышечных сокращений мочеточников, но и тонусом мускулатуры мочевого пузыря. Каждому поступлению мочи из мочеточников предшествует дилятация мочевого пузыря, создаваемая растяжением его мышц. В результате этого возникает снижение давления в мочевом пузыре, что наряду с сокращением мускулатуры мочеточника способствует свободному поступлению мочи в мочевой пузырь. Этот процесс по аналогии можно сравнить с поступлением мочи из почечного сосочка в чашечку. Иначе говоря, растяжение стенок мочевого пузыря создает снижение давления в нем, что обеспечивает „отсасывание" мочи из мочеточников наряду с мышечным сокращением нижних цистоидов. После опорожнения нижних цистоидов происходит падение давления в них, в результате чего возникает спадение стенок мочеточника, что в комплексе с другими моментами (см. „Физиология верхних мочевых путей") препятствует обратному проникновению мочи, т. е. развитию пузырно-мочеточникового рефлюкса.
Таким образом, фаза диастолы мочевого пузыря есть не пассивный, а активный акт, в котором активно участвует детрузор. Подтверждением активной деятельности детрузора и его большой роли в обеспечении нормального пассажа мочи из верхних мочевых путей могут служить данные цистометрии. Давление мочи в мочевом пузыре в состоянии так называемого покоя равно 5—10 см вод. ст., и нет значительных отклонений по мере наполнения мочевого пузыря. Однако налицо колебания давления в указанных пределах. Снижение давления есть результат растяжения детрузора и увеличения объема пузыря, которые являются как бы подготовительной фазой для поступления мочи из мочеточников. После того как очередная порция мочи поступит в мочевой пузырь, возникает подъем давления. Этот подъем давления есть результат не только увеличения объема мочевого пузыря, а преимущественно усиления тонуса детрузора. Вслед за этим возникает вновь снижение давления в мочевом пузыре. И так происходит до достижения нормально допустимой емкости мочевого пузыря. Эти данные цистометрии подтверждают активную функцию мочевого пузыря в фазе диастолы, чем и обеспечивается нормальный пассаж мочи из мочеточников.
Функция верхних мочевых путей и в первую очередь мочеточников теснейшим образом связана с функцией мочевого пузыря. В основе этой функциональной взаимосвязи лежит рефлекторный процесс, усиление и ускорение деятельности мочевого пузыря в его диастолической фазе. Перерастяжение мочевого пузыря и повышение в нем давления выше нормального — предельно допустимого (25—40 см вод. ст.) — вызывает резкое замедление динамики опорожнения мочеточников вплоть до полного его прекращения (адинамия, акинезия, гипотония). Это может быть иллюстрировано следующим примером. Если хромоцистоскопия выполняется при переполненном мочевом пузыре, то наблюдается резкое торможение динамики опорожнения мочеточников; об этом можно судить по прекращению сокращений мочеточниковых устий и отсутствию своевременного выделения индигокармина, несмотря на то что функциональная способность верхних мочевых путей и мочевого пузыря не нарушена. Подобные явления можно наблюдать и в тех случаях, когда цистоскопией вызвана даже незначительная травма задней уретры и шейки мочевого пузыря. Возникшее в результате этого торможение функции пузыря влечет за собой торможение функции мочеточников. Эта теснейшая рефлекторная взаимосвязь в деятельности мочевого пузыря и верхних мочевых путей положена в основу экскреторной урографии в комбинации с кислородной цис-тографией по А. Я. Пытелю, когда рефлекс с переполненного газом мочевого пузыря вызывает замедление динамики опорожнения верхних мочевых путей, повышение в них давления, следствием чего является активная функция форникального аппарата почки. Данный факт является доказательством рефлекторной связи деятельности мочевого пузыря не только с мочеточниками, но и с почечной лоханкой, чашечками и их форникаль-ными зонами. Достаточно повысить давление в мочевом пузыре до 15 см вод. ст., как резко нарушается уродинамика мочеточников, почечных лоханок и их чашечек. В основе этих рефлекторных реакций лежит не только барорецепция. но в большой степени механорецепция; окончания баро-и механорецепторов реагируют при изменении тонуса детрузора, которое возникает вследствие растяжения стенок мочевого пузыря.
Таким образом, функция мочевого пузыря теснейшим образом связана с функцией верхних мочевых путей и наоборот. Только согласованная деятельность всех звеньев мочевыводящих путей, регулируемая нервной системой, может обеспечить весь сложный процесс уродинамики — эвакуации мочи из организма.
Литература
Баялова С. А. Труды Волгоградск. мед. ин-та, 1951, 8, 297.
Баялова С. А. Морфология гладкой мускулатуры верхних мочевых путей. Дисс.
Сталинград, 1952.
Пытель А. Я. Хирургия, 1950, 7, 8.
Пытель А. Я. Лоханочно-почечные рефлюксы и их клиническое значение. М., 1959. Пытель Ю. А. Урология, 1955, 3, 50. Пытель Ю. А. Урология, I960, б, 9. X а ю т и н В. М. Мочеиспускание. БМЭ, 1961, т. 19, 259.
В е g g R. С. Brit. J. UroL, 1946, 18, 4, 176.
BodnerH., Howard A. H a. Kaplan J. H. J UroL, 1958, 79, 356.
EmmettJ. L. Physiology of the normal bladder. In: Campbell M. F., Urology. Philadelphia, 1954.
F e у В., H e n i F., Kuntz A., McDonald D. F., QuenuL., Wesson L. G., Wilson G. Physiology and pathological physiology. Encvcilopeda of Urology. Vol. II. Berlin, 1964.
F e у В., Т г и с h о t P., N о i x M. XI Congress Internat. Urologie. Stockholm, 1958, p. 138.
F e у В., Т г и с h о t P. et N о i x M. J. Radiol. Electrol., 1958, 39, 328.
FuchsF. Ztschr. UroL Chir., 1931, 33, 1.
FuchsF.J. UroL, 1934, 32, 115.
H a n 1 e у H. G. Proc. roy. Soc. Med., 1956, 49, 342.
H a n 1 e у H. G. Brit. J. UroL, 1957, 29, 312.
H u t с h J. A. Invest. UroL, 1965, 3, 36.
К i i 1 F. The function of the ureter and renal pelvis. Philadelphia, 1957.
L a p i d e s J. J. UroL 1948, 59, 501.
L a p i d e s J., H о d s о n N. В., В о у d R. E., Snook E. L. a. Lichtwardt I. J. UroL, 1958., 79, 4, 707.
LongworthyO. R.,KolbL. С a. Lewis L. G. Physiology of Micturition. Baltimore, 1940.
M i t s u у a H., A s a i J., S u у a m a K., S a i E. a. H о s о е К. UroL Internat., 1962, 13, 236.
N a r a t h P. A. Renal pelvis and ureter. New York, 1951.
Renyi-VamosF.,BaloghF. a. SzendroiZ. ActaUrol. (Budapest), 1948,2,103.
Saccomanso (1944). Cit. J. L. Emmett, 1954.
Schneider W. Ztschr. Anat. u. EntwickL, 1938, 109, 2, 187.
S u у a m a K. Japan. J. UroL, 1961, 52, 463.
T a n a g h о Е. a. PughR. Brit. J. UroL, 1963, 35, 151.
Глава пятая
Физиология
мужских половых органов
Внутренние половые органы
Яички обладают двойной функцией: генеративной, которая служит для выработки сперматозоидов и поддержания рода, и внутрисекреторной, которая заключается в образовании специфического гормона, способствующего проявлению вторичных половых признаков и нормального полового влечения и стимулирующим образом действующего на нервно-мышечную систему организма.
Сперматогенная функция связана с зародышевым эпителием извитых канальцев. Собственная оболочка извитых канальцев выстлана клетками двоякого рода: фолликулярным эпителием, или клетками Сертоли, и половыми, или семенными, клетками (сперматогоыии), развивающимися из первичных зародышевых клеток в период полового созревания. Клетки Сертоли прилежат своим основанием к базальной мембране канальца, а тело в виде цилиндрической колонки проходит через всю стенку канальца до самого просвета; они являются питающими сперматозоиды клетками.
Из сперматогоний, лежащих у стенки канальца на расширенных ножках клеток Сертоли, образуются сперматоциты первого порядка, а из них в свою очередь — сперматоциты второго порядка (пресперматиды). Пре-сперматиды образуют сперматиды, которые развиваются в сперматозоиды (рис. 32).
Процесс формирования сперматозоидов происходит в клетках Сертоли. К моменту возникновения сперматид в клетках Сертоли образуются протоплазменные выросты, которые поглощают сперматиды. Развивающиеся из сперматидов сперматозоиды погружены своими головками в протоплазменные выросты клеток Сертоли, а их хвосты обращены в просвет канальца; в каждой клетке находится 8—12 сперматозоидов. После распада протоплазменных выростов сперматозоиды освобождаются. Продолжительность образования сперматогоний в зрелые сперматозоиды равна в среднем 19—20 дням.
Выработка сперматозоидов начинается в период полового созревания (13—17 лет) и происходит непрерывно до старости. Начало этого периода подвержено довольно значительным индивидуальным колебаниям. Самым ранним сроком появления эякулята, содержащего сперматозоиды, следует считать 13-й год (Rohleder, 1925; Fiirbringer, 1890). Однако Simmonds (1898) исследовал 1000 трупов мальчиков, умерших до 15-летнего возраста, и ни разу не нашел сперматозоидов.
Согласно гистологическим исследованиям Stieve (1930) и Т. С. Ломовиц-кой (1958), периодом морфологического и функционального развития зародышевых желез следует считать возраст от 14 до 15 лет. Однако зрелые сперматозоиды в яичках появляются лишь в возрасте 16 лет (Stieve, 1930; М. А. Лейзеруков, 1964).
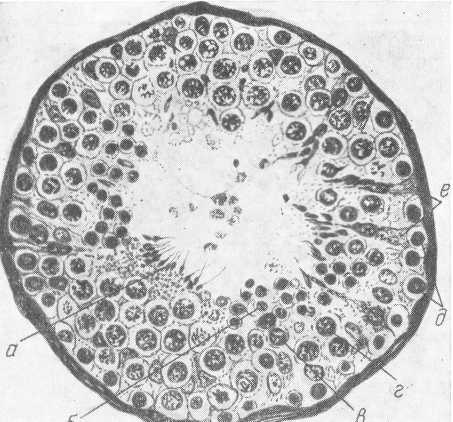
Рис. 32. Поперечный разрез канальца яичка (по Штиве).
а — сперматозоиды; б — сперматиды; в — пресперматиды; г — сперматоциды;
д — сперматогонии; е — клетки Сертоли.
Что касается вопроса об угасании сперматогенеза в старости, то он вляяется малоизученным, так как в глубокой старости отсутствует возможность доказательства способности к оплодотворению. Simmonds (1898) не признает никаких возрастных ограничений для сперматогенной функции яичек. Согласно данным Rohleder (1925), сперматозоиды в эякуляте обнаруживаются в возрасте до 70 лет в большинстве случаев, в возрасте 70—80 лет — в половине, в возрасте 80 — 90 лет — в четверти и в возрасте старше 90 — в незначительном количестве случаев. Belonoschkin (1949, 1954) из 83 обследованных мужчин в возрасте 60—92 лет у 2/з из них. обнаружил живые сперматозоиды. Известный интерес представляют патолого-анатомические данные Dyplae (1852) и Dien (1867), которые при исследовании 165 трупов стариков обнаружили сперматозоиды у 68,5% умерших в возрасте 60—70 лет; у 59,8% — в возрасте 70—80 лет и у 48 % — в возрасте 80—90 лет. Seymour, Duffy, Kerner (1935) сообщили о способности к оплодотворению 94-летнего мужчины. Doepfmer (1957) наблюдал 74-летнего мужчину, эякулят которого содержал 163 млн. сперматозоидов в 1 мл. Мы наблюдали 68-летнего мужчину, от которого женщина в течение l1 лет два раза забеременела. В эякуляте у него при микроскопическом исследовании все поле зрения было полно сперматозоидов, большинство которых находршись в состоянии активной подвижности. Заслуживает внимания такой факт: в 1955 г. в Западной Германии, согласно данным статистического управления, родилось около 200 детей от мужчин в возрасте 66 лет и старше (Nikolowski 1960).
Однако следует указать, что в старости сперматогеыная функция и способность к оплодотворению постепенно регрессируют. Это стоит в связи с теми структурными изменениями, которые наступают в яичках в старости. С возрастом постепенно уменьшается количество клеток Лейдига, развиваются явления перитубулярного, а затем и тубулярного склероза с облитерацией канальцев (Tillinger, 1957; М. А. Лейзеруков, 1964, и др.). В результате прогрессирующего ухудшения кровообращения и питания яичка значительно падает активность сперматогенной функции. В связи с этим уменьшается количество сперматозоидов в эякуляте и нарастает количество неподвижных и патологических форм. Физиологическое падение гормональной функции яичек ведет к нарушению секреторной функции добавочных половых желез. Это влечет за собой уменьшение образования фруктозы, что в свою очередь оказывает неблагоприятное влияние на жизнеспособность и подвижность сперматозоидов. Существуют, однако, выраженные индивидуальные колебания в отношении срока угасания сперматогенеза и способности к оплодотворению у мужчин. Сохранение этой способности в старости не является, как указано выше, редкостью. Очевидно, в старости сперматогенная функция сохраняется значительно дольше, чем гормональная. Если, однако, у женщины остановка созревания фолликулов в яичнике и связанное с этим прекращение менструаций (менопауза) происходят в относительно ограниченный период жизни, в среднем от 45 до 50 лет, то у мужчин не существует строго определенного возраста, когда сперматогенная функция полностью потухает.
Внутрисекреторная функция яичка сосредоточена в межуточных клетках Лейдига. Исследования последнего времени дали новые доказательства того, что половой гормон (тестостерон) образуется именно в клетках Лейдига. Rohmeis (1943) производил кастрированным котам внутрискротально аутопластические прививки кусочков ткани яичка, состоящих исключительно из межуточных клеток. Кастрированные животные полностью сохраняли при этом свои вторичные половые признаки. Не меньшее значение имеют исследования Hooker (1948), которому удалось установить тесную связь между содержанием в яичке межуточных клеток и гормонов, а также Pollok (1942), доказавшего, что стероидные вещества с химическими свойствами тестостерона содержатся только в межуточных клетках, а не в каких-либо других составных частях яичка.
Установлено также, что в яичке человека имеет место образование эстрогена; при этом речь идет об эстрадиоле, продукция которого происходит в межуточных клетках.
С повышением возраста происходит понижение гормональной функции клеток Лейдига (Hamilton H. и Hamilton J., 1948; Hamburger, 1951).
В связи с выпадением тормозящего влияния яичек на переднюю долю гипофиза в старческом возрасте наступает увеличенное выделение гонадо-тропного гормона.
В старости происходит падение не только функции яичек, но и деятельности предстательной железы. После 50 лет уменьшается количество железистой ткани, эпителиальные клетки становятся более низкими, кубическими, начинает преобладать фиброзпо-мышечная ткань, количество липоидных зерен, являющихся секреторным продуктом железистого эпителия предстательной железы, после 50 лет уменьшается (И. М. Порудоминский и М. Б. Фейгин, 1930). В зрелые годы отмечается более или менее значительное падение содержания фосфатазы в эякуляте, которая образуется в предстательной железе. Аналогичное явление имеет место с содержанием фруктозы в эякуляте, продукция которой происходит в семенных пузырьках. С повышением возраста постепенно падает содержание фруктозы в семенной плазме.
Придаток яичка является секреторным органом, имеющим несомненное физиологическое значение для оплодотворяющей способности мужчины. Выработка секрета придатка поддерживается тестикулярным гормоном (Moore, 1935; Б. С. Гехман, 1958, 1963, и др.).
Придаток является резервуаром для сперматозоидов. Концентрация сперматозоидов возрастает от яичка и головки придатка к хвосту. После повторных электрокоагуляций количество сперматозоидов в головке и теле не изменяется, в то время как в хвосте придатка оно постепенно уменьшается. Головка и тело придатка являются, по-видимому, лишь отрезками, по которым передвигаются сперматозоиды, в то время как накопление их происходит в хвосте (Ortavant). Поэтому именно хвост придатка рассматривается как резервуар для накопления смерматозоидов (Boeminghaus, 1955; Korkud, 1955; А. В. Немилов, 1933; В. В. Пловцева, 1937; Б. С. Гехман, 1963, и др.). В придатке яичка происходит дальнейшее морфологическое и физиологическое развитие сперматозоидов.
Сперматозоиды, которые прошли через придаток, обладают более интенсивной подвижностью и жизнеспособностью и в отличие от сперматозоидов яичка приобретают способность к оплодотворению. Оплодотворяющая способность сперматозоидов увеличивается по мере продвижения от головки к хвосту придатка: 32% оплодотворения могут быть получены сперматозоидами головки и 68% сперматозоидами хвоста (Young, 1931).
Семенные пузырьки являются железистыми органами, которые развиваются и функционируют иод влиянием андрогена. После кастрации эпителий семенных пузырьков становится низким, секреторная деятельность его уменьшается. После введения тестостерона атрофированный эпителий вновь восстанавливается до нормы, при этом в тесной зависимости от величины введенной дозы и длительности применения. Эта зависимость настолько отчетливо выражена, что используется как тест для количественного определения андрогена.
Секрет семенных пузырьков имеет серовато-белый цвет, клейкую, же-латиноиодобную консистенцию. Выделяясь наружу, он приобретает вид саговых зерен, быстро растворяющихся в эякуляте. Концентрация водородных ионов (рН) секрета равна 7,3. Секрет семенных пузырьков вместе с секретом предстательной железы составляет большую часть семенной жидкости. Наиболее важной составной частью секрета семенных пузырьков является фруктоза. Выделение фруктозы стимулируется и регулируется половым гормоном. Уровень фруктозы является показателем внутрисекреторной функции зародышевых желез.
Определение фруктозы производится в фильтрате семени, освобожденном от белков посредством сульфата цинка и едкого натра на основании колориметрической реакции фруктозы с резерцином. Интенсивность окраски прямо пропорциональна концентрации фруктозы. Метод определения фруктозы разработан Roe (1934), в основу его положена цветная реакция, открытая Seliwanoff (1887). Нормальное содержание фруктозы, согласно данным Nowakowski и Schirren (1956), Gropper и Nikolowski (1954) и др., находится между 1200 и 3500 у/мл семенной жидкости; согласно данным Kimmig (1950), — между 1200 и 4000 у/мл, а по Wasterling (1959) — между 1000 и 5000 у/мл. Среднее количество фруктозы, по данным Wasterling, Davis, McCune (1950), Eichenberger и Goosens (1950), Gropper и Nikolowski (1954), Nowakowski и Schirren (1956), составляет 290 мг%, по данным М. А. Кунина (1963), — 250 мг%. Количество фруктозы ниже физиологической нормы при отсутствии патологических изменений в семенных пузырьках говорит о понижении функции клеток Лейдига.
Фруктоза является тем субстратом, который необходим сперматозоидам для процессов обмена веществ (Mann, 1948). Она представляет собой существенное, но не исключительное средство, способствующее проявлению подвижности сперматозоидов. Только нормальные сперматозоиды в состоянии использовать имеющуюся в семенной плазме фруктозу; патологические сперматозоиды и тогда остаются неподвижными, когда в семенной плазме находится много фруктозы. Недостаток фруктозы может повлечь за собой понижение подвижности сперматозоидов. Быстрота расщепления фруктозы (фруктолиз) была предложена Mann для определения качества эякулята при искусственном оплодотворении, в котором фруктоза играет большую роль. На основании фруктолиза можно судить о подвижности и жизнеспособности сперматозоидов.
Секрет семенных пузырьков содержит также близко стоящий к сахарам циклический алкоголь инозит. Schirren (1961) считает, что инозит образуется в простате. Инозит существует в виде 8 изомеров, из которых в семен-
ной плазме находится мезоинозит. Значение инозита в семени остается неизвестным. На подвижность сперматозоидов и на фруктолиз он не оказывает влияния. У больных с эпидимитом и везикулитом обнаруживают уменьшение количества инозита, что связано с понижением синтеза его в результате воспалительного процесса в семенных пузырьках.
Количественное содержание инозита в семени определяется микробиологическим методом, который основан на измерении роста дрожжевых грибов типа Юоссега brevis. Скорость роста дрожжей пропорциональна количеству инозита; оценка производится турбидиметрическим методом (по степени мутности субстрата). Содержание инозита в человеческом семени колеблется, по данным разных авторов, от 114 до 2500 у/мл (Kimmig и Schirren, 1956). В норме содержание инозита колеблется от 1000 до 2500 у/мл (Schirren, 1961).
Секрет семенных пузырьков, примешиваясь к эякуляту, придает ему вязкость и студенистость, способствует увеличению его массы, что имеет значение для оплодотворения.
В семенных пузырьках всегда находят более или менее значительное количество сперматозоидов. Они попадают в семенные пузырьки из ампул семявыносящего протока в случае их чрезмерного наполнения сперматозоидами. Они проникают в семенные пузырьки также при эротическом возбуждении без последующей эякуляции, как это было доказано экспериментальными исследованиями. В настоящее время следует считать установленным, что семенные пузырьки не являются резервуаром для сперматозоидов, как это было принято в свое время считать и как это думают еще многие врачи в настоящее время. Резервуаром для сперматозоидов, как уже было сказано, является хвост придатка, в котором всегда находится значительное количество сперматозоидов.
Предстательная железа выделяет секрет слабо щелочной реакции, богатый протеином. В нем содержится большое количество преломляющих свет зернышек, которые являются специфическим секреторным продуктом железистого эпителия предстательной железы. Указанные зернышки придают секрету простаты молочный вид. Они принадлежат к липоидам в широком смысле слова и относятся к группе фосфатидов, к каковым относится лецитин (И. М. Порудоминский и М. Б. Фейгин, 1930). В секрете простаты содержится спермин, который сообщает семени его характерный запах. В эякуляте он существует не в свободном виде, а в виде фосфата спермина. Кристаллы фосфорного спермина образуются только после эякуляции в результате соединения спермина с фосфорной кислотой, освобождающейся после расщепления холинофосфорной кислоты под влиянием фосфатазы (см. ниже).
О значении спермина, а также содержащегося в семенной плазме спер-мидина ничего определенного сказать нельзя. Они, по-видимому, способствуют поддержанию на соответствующем уровне концентрации водородных ионов.
Содержание спермина колеблется в пределах 254—3180 у/мл. Средняя концентрация спермина равна 1320 у/мл (Schirren).
Секрет простаты содержит также два фермента — фибролизин и фибро-геназу (Huggens и Neal, 1942). Они принимают участие в разжижении семени и переводят в жидкое состояние свернувшийся эякулят. Фибролизин является весьма сильным протеолитическим ферментом; 2 мл этого фермента в состоянии разжижать 100 мл человеческой коагулированной крови. Количество фибролизина у человека больше фиброгеыазы.
В секрете предстательной железы имеется в значительном количестве лимонная кислота. Содержание лимонной кислоты в секрете простаты колеблется от 480 до 2688 мг% (Huggens и Neal, 1942).
В среднем в человеческом семени содержится 479 мг% лимонной кислоты (Harvey, 1951).
Лимонная кислота в секрете простаты появляется с наступлением половой зрелости, исчезает после кастрации и вновь возвращается к нормальной величине после введения тестостерона (Mann, 1953; Litwak-Mann и Mann, 1951; Schirren, 1961, и др.). Зависимость между образованием лимонной кислоты и содержанием тестостерона настолько отчетливо выражена, что можно по количеству лимонной кислоты в секрете простаты судить о функциональном состоянии клеток Лейдига.
Значение лимонной кислоты в предстательной железе и семенной плазме еще недостаточно выяснено. Можно сделать предположение, что она имеет отношение к свертыванию и разжижению семени, равно и к образованию ионов кальция и активации гиалуронидазы (Kimmig, 1955). Определение лимонной кислоты в семени можно использовать для выяснения функционального состояния предстательной железы.
Определение лимонной кислоты производится в фильтрате семени, освобожденном от белков. Оно основано на окислении лимонной кислоты в пентобромацетат с последующей экстракцией петролейным эфиром и оценивается колориметрически.
Секрет предстательной железы является одним из наиболее богатых источников кислой фосфатазы в человеческом организме. В простате имеется также и щелочная фосфатаза, но количество ее невелико. Образование кислой фосфатазы в простате, как и лимонной кислоты, тесно связано с инкреторной функцией зародышевых желез, оно резко повышается при половом созревании и угасает с прекращением образования тестостерона в яичках. Эстроген вызывает снижение кислой фосфатазы в сыворотке подобно кастрации. Применение андрогена, наоборот, вызывает значительное ее нарастание.
Рак предстательной железы, в особенности при метастазах, сопровождается, как известно, повышенным образованием кислой фосфатазы. Поэтому исследование крови на кислую фосфатазу имеет известное значение для диагностики этого заболевания.
Фосфатаза простаты играет, по-видимому, роль в процессе оплодотворения, тем не менее нельзя привести еще какие-либо достоверные данные в пользу этого. Она, возможно, принимает участие в углеводном обмене сперматозоидов (Mann). Несомненным является то, что под действием фосфатазы происходит расщепление холинофосфорной кислоты семенной плазмы на холин и фосфорную кислоту. В течение 20 минут после эякуляции количество фосфорной кислоты повышается от 10 до 60 мг% (Kimmig, 1955).
В результате этого происходит значительное обогащение эякулята свободным фосфором и холином. Согласно исследованиям Werner и Becker, холин оказывает сенсибилизирующее влияние на клетки. Так, например, опухолевые клетки под влиянием холина становятся более чувствительными к рентгеновым лучам. Для их разрушения требуются дозы в 2 раза меньшие, чем для опухолевых клеток, не обогащенных холином. Таким образом, возможно, что фосфатаза оказывает косвенное сенсибилизирующее влияние на сперматозоиды. Наиболее удобным методом определения фосфатазы является метод Кирбергера и Мартини (Kirberger и Martini, 1950). Количественное содержание фосфатазы вычисляется по количеству образованного фосфорпентоксида в единицу времени (Р2О5/ЗО минут). Нормальное содержание фосфатазы в предстательной железе составляет 100—300 Р2О5/30 минут.
Приведенные данные позволяют считать, что секрет предстательной железы участвует в разжижении семени, увеличении его объема, а также
в буферном и ферментативном влиянии семени, в то время как секрет семенных пузырьков служит для метаболизма сперматозоидов и создает условия для повышения их подвижности и жизнеспособности.
Многими авторами высказывалось предположение об эндокринной функции предстательной железы. Однако убедительные данные в пользу этого до сих пор отсутствуют.
Существуют весьма тесные взаимоотношения между предстательной железой и яичками. Опыты на гипофизэктомированных животных (крысы, собаки, морские свинки, резус-обезьяны и др.) показали, что введение экстракта яичек задерживает наступление атрофии простаты и вызывает увеличение ее веса у старых животных. Аналогичное действие оказывает введение тестостерона, при этом особенно стимулируется развитие железистого эпителия предстательной жезезы и максимально нарастает его секреторная деятельность. Клинические наблюдения над людьми также показали, что кастрация вызывает изменения в росте и развитии предстательной железы. В зрелом возрасте кастрация сопровождается атрофией предстательной железы. Кастрация же, произведенная тогда, когда предстательная железа еще не достигла полного развития, вызывает остановку ее роста. Развитие и деятельность предстательной железы находятся, таким образом, под влиянием андрогенной функции яичек.
Предстательная железа оказывает влияние на функцию яичек. Согласно опытам Serralach и Pares (1911), экстирпация предстательной железы у собак сопровождается атрофией яичек и исчезновением сперматозоидов в семени. Впрыскивание экстракта предстательной железы влияет на сперматогенез: малые дозы экстракта возбуждают, большие — угнетают сперматогенез. Наблюдения других авторов также показали, что половые органы у собаки, кролика и морской свинки развиваются после применения экстракта простаты скорее и половое созревание наступает преждевременно.
Н. А. Белов (1912) считает, что яички оказывают на предстательную железу прямое действие, а предстательная железа на яички — перекрестное действие. Деятельность предстательной железы возбуждается при усиленной функции яичек и угнетается при понижении их функции. Наоборот, усиленное всасывание в кровь секрета предстательной железы угнетает деятельность яичек, в то время как уменьшение всасывания секрета возбуждает их функцию.
Известно, например, что чем чаще совершается половой акт, тем сильнее стремление к нему. Это можно объяснить тем, что при активной половой деятельности большие количества секрета предстательной железы выбрасываются наружу, в результате чего в кровь поступают малые дозы его, что приводит к усилению функции яичек и повышению полового влечения. Наоборот, чем реже совершается половой акт, тем меньше стремление к нему, так как поступающий в больших количествах в кровь секрет предстательной железы тормозит деятельность яичек.
Не умаляя значения приведенных исследований, следует, однако, сказать, что они не являются достаточными для заключения о внутрисекреторной деятельности предстательной железы.
Секрет предстательной железы способствует разжижению семени и увеличению его объема. Он активирует движение сперматозоидов и создает условия для повышения их жизнеспособности. Это свойство секрета простаты не является, однако, обязательным для оплодотворения, так как последнее, как уже было указано, может быть достигнуто сперматозоидами, добытыми из хвоста придатка.
Тесная эмбриологическая и топографическая связь предстательной железы с семявыводящими путями, зависимость ее развития и функционирования железы от половых гормонов позволяют думать, что предстательная железа принимает участие в половой функции.
Каким путем осуществляется влияние предстательной железы на половую способность, нельзя считать окончательно изученным. Можно думать, что оно происходит рефлекторным путем через периферические нервные окончания. Патологические изменения в предстательной железе, вызывая нарушение чувствительности рецепторного аппарата (повышение или понижение возбудимости), могут привести к различным расстройствам половой деятельности. Качественные различия в изменении возбудимости рецепторного аппарата зависят от характера и особенностей патологических процессов в предстательной железе.
Значение заболеваний предстательной железы в этиологии половых расстройств подтверждается многочисленными клиническими наблюдениями.
Наружные половые органы
Мошонка обладает терморегулирующей функцией. Температура в мошонке на 3—4° ниже, чем в брюшной полости, что имеет большое значение для нормального развития и созревания сперматогенных клеток. В тех случаях, когда яичко не опускается в мошонку, а задерживается в брюшной полости или в паховом канале, сперматогенез резко нарушается. Межуточные клетки при этом мало страдают. Механические раздражения вызывают сокращения мускула кремастера (кремасторный рефлекс), а холод — сокращение мясистой оболочки мошонки. В обоих случаях яичко приподнимается кверху, к более теплому туловищу. Наоборот, сильное нагревание вызывает расслабление тканей и отодвигание яичка от туловища.
Половой член предназначен для совершения полового акта, чему способствует его анатомическое строение (наличие кавернозной ткани). В обычных условиях при расслабленном состоянии полового члена сосуды находятся в тоническом сокращении. Если этот тонус внезапно исчезает, то кровь устремляется в значительно расширившиеся сосуды и их конечные резервуары — каверны, что приводит к увеличению объема полового члена и его резкому уплотнению. Этому в значительной мере способствует также сдав-ление расширенными кавернами венозных сосудов и затруднение оттока крови из кавернозных тел, окруженных мощными белочными оболочками. Некоторое подсобное значение, возможно, имеет и сокращение луковично-пещеристой и седалищно-пещеристой мышц, способствующее более энергичному проталкиванию крови в тело полового члена и усиливающее тем самым эрекцию. Однако главное назначение луковично-пещеристой мышцы в том, чтобы обеспечить в момент оргазма выбрасывание семени из уретры.
Кавернозное тело уретры имеет несколько иное строение. Вследствие менее мощной белочной оболочки отток крови из кавернозного тела уретры во время эрекции не столь затруднен, как из кавернозных тел полового члена. В результате этого даже на высоте эрекции кавернозное тело уретры и головка члена остаются менее напряженными и более податливыми; этим самым создаются благоприятные условия для прохождения семени через уретру.
Известную роль в половой функции играют куперовские и литтреевские железы уретры, секреты этих желез благодаря своей щелочной реакции нейтрализуют кислую реакцию уретры, возникающую после мочеиспускания. Кроме того, смазывание слизистой оболочки уретры этими секретами облегчает прохождение эякулята во время полового акта.
Расположенный в задней уретре семенной бугорок принимает, очевидно, участие в эякуляционной функции (А. И. Васильев, 1913; Р. М. Фронштейн, 1938; Л. Я. Якобзон, 1915). В пользу этого говорит ряд факторов: эмбриологическая связь, которая существует между органами, принимающими участие в эякуляции (предстательная железа, семенные пузырьки и семенной бугорок развиваются из первичной матки), концентрация выводных протоков этих органов вокруг семенного бугорка, а также то, что при задней уретроскопии нередко удается наблюдать выбрасывание семени из устьев семявыносящих протоков, вызванное механическим раздражением семенного бугорка инструментом. Кроме того, клинические наблюдения показывают, что воспалительные изменения в задней уретре и семенном бугорке вызывают нередко поллюции и преждевременное семяизвержение при половом акте. Приведенные данные позволяют считать, что семенной бугорок является местом концентраций нервных окончаний афферентных путей, участвующих, в акте эякуляции.
Нейро-эндокринная регуляция
половой функции
ВЗАИМОСВЯЗЬ МЕЖДУ СЕМЕННЫМИ
И ДРУГИМИ ЭНДОКРИННЫМИ ЖЕЛЕЗАМИ
Главным источником половых гормонов у мужчин являются яички. Функция яичек находится в тесной коррелятивной связи с функцией других органов внутренней секреции: гипофизом, надпочечниками, щитовидной и зобной железами (рис. 33). Наибольшее значение имеет коррелятивная связь, существующая между яичками и гипофизом. Известно, что передняя доля гипофиза выделяет 2 гонадотропных гормона: 1) гонадотропный фол-ликулостимулирующий гормон (FSH); он же обладает специфическим стимулирующим действием на сперматогенный эпителий, побуждая его к выработке сперматозоидов, и 2) гонадотропный лютеинизирующий гормон, стимулирующий клетки межуточной (интерстициальной) ткани (ICSH); он же оказывает стимулирующее влияние на клетки Лейдига, побуждая их к выработке полового гормона — тестостерона.
Гонадотропный гормон передней доли гипофиза оказывает чрезвычайно большое влияние на развитие и нормальную функцию яичек, являясь активным стимулятором („мотором") половой функции. После удаления гипофиза задерживается развитие семенников и вторичных половых признаков.
Влияние гипофиза является определяющим для образования андрогена, для развития и сохранения сперматогенеза. Однако влияние гипофиза на сперматогенез не является единственным.
Для образования сперматозоидов необходимо существование нормально функционирующих клеток Лейдига. Сперматогенез находится под влиянием фолликулостимулирующего гормона (FSH) гипофиза и тестостерона, который вырабатывается клетками Лейдига яичка, находящимися в тесном контакте с семенными канальцами. Тестостерон непосредственно, а не через кровеносные пути способствует развитию и сохранению зародышевого эпителия канальцев. Так как анатомические субстраты внутренней и внешней секреции яичка находятся в непосредственной близости друг от друга, то это приводит к тому, что в данном органе сохраняется необходимый для нормальной жизнедеятельности сперматозоидов высокий уровень гормона. Тестостерон оказывает важное трофическое влияние на семенные канальцы (Tonnuti, 1955).
Рис. 33. Гормональные взаимоотношения между передней долей гипофиза,