

1409
.pdfУДК 531/524:[57+61]
Е.В. Феоктистова, М.С. Куюкина, М.А. Осипенко
E.V. Feoktistova, M.S. Kuyukina, M.A. Osipenko
Пермский национальный исследовательский политехнический университет
Perm National Research Polytechnic University
БИОМЕХАНИЧЕСКАЯ МОДЕЛЬ МЕЖФАЗНЫХ ПРОЦЕССОВ В СИСТЕМЕ БАКТЕРИИ – ВОДА – УГЛЕВОДОРОД
BIOMECHANICAL MODEL OF INTERFACIAL PROCESSES IN THE SYSTEM BACTERIA–WATER–HYDROCARBON
Рассмотрены поверхностные явления на межфазной границе, разделяющей клеточную суспензию бактерий рода Rhodococcus и жидкий углеводород. Проведено детерминистическое и вероятностное моделирование временной зависимости коэффициента поверхностного натяжения. Нестабильный характер зависимости объяснен случайными изменениями степени образования единой пленки бактерий.
The surface phenomena on the interface dividing the suspension of Rhodococcus cells and the liquid hydrocarbon are considered. The deterministic and the stochastic modelling of the time dependence of surface tension coefficient are carried out. The instability of this dependence is explained by the random variations of the degree of the formation of entire film of cells.
Ключевые слова: бактериальные клетки, Rhodococcus, поверхностное натяжение, детерминистическая модель, вероятностная модель.
Keywords: bacterial cells, Rhodococcus, surface tension, deterministic model, stochastic model.
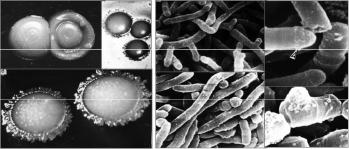
Актинобактерии рода Rhodococcus (рис. 1) являются представителями эко- лого-трофической группы микроорганизмов, окисляющих природные и антропогенные углеводороды и, следовательно, участвующих в различных биогеохимических процессахиформированиибезуглеводороднойбиосферы[1].
Родококки способны к окислению углеводородов нефти, смол, фенольных и полихлорированных соединений, гумусовых веществ, лигнина и его производных, восков, пестицидов и др. Некоторые родококки способны усваивать непредельные углеводородные соединения, например ацетилен ипропен, а также ароматические– фенолы, хлорфенолы, ароматическиеуглеводородынефти.
Реализация биотехнологического потенциала родококков предусматривает всестороннее изучение механизмов их адгезии к различным веществам. В частности, в лаборатории алканотрофных микроорганизмов Института экологии и генетики микроорганизмов УрО РАН изучаются закономерности адгезии клеток родококков к жидким алканам. Межфазное натяжение на грани-
221
це раздела клеточной суспензии и углеводорода измеряется с помощью уста-
новки Sigma 701.
Рис. 1. Колонии (а) и клетки (б) актинобактерий рода Rhodococcus
Анализ экспериментальных данных. На рис. 2 представлены результа-
ты долгосрочных экспериментов по изучению динамики межфазного натяжения различных концентраций клеток в системе вода-н-гексадекан. Установлено, что после 6 ч взаимодействия клеток с н-гексадеканом наблюдается скачкообразный характер кривой межфазного натяжения при средних (0,75–1,0) значениях оптической плотности ОП600нм (ОП – безразмерная величина, характеризующая степень погашения света, прошедшего через слой материала), что, по-видимому, свидетельствует о постепенной стабилизации процесса адсорбции клеток в межфазном слое. При определении показателей межфазного натяжения клеточной суспензии в концентрации ОП600нм = 1,25 стабильных значений в течение 8 ч эксперимента зафиксировать не удалось.
Установлено, что после 9 ч взаимодействия клеток с н-гексадеканом показатели межфазного натяжения приобретают стабильный характер (28–29 мН/м), что говорит о предельном насыщении межфазного слоя клетками родококков и образовании межклеточных контактов.
Для исследования влияния природы гидрофобного субстрата на динамику процесса адгезии клеток в двухфазной системе была проведена серия экспериментов с н-додеканом (рис. 3), обладающим меньшей гидрофобностью, чем н-гексадекан, и, соответственно, большей растворимостью в воде.
В результате проведенных исследований показано, что углеводородный субстрат оказывает влияние на динамику межфазного натяжения. Как видно из рис. 3, в системе с н-додеканом к 19 ч эксперимента показатели межфазного натяжения достигают постоянной величины (27 мН/м) уже при минимальной концентрации клеточной суспензии (ОП600нм = 0,25). Интересно, что при использовании суспензии клеток с концентрацией ОП600нм = 0,5, стабильных показателей межфазного натяжения зафиксировать не удалось, т.е. данная концентрация клеток является переходной к стабильной системе. Следует отметить, что стабильные показатели межфазного натяжения регистрируются уже при средней концентрации клеток (ОП600нм = 1,25).
222
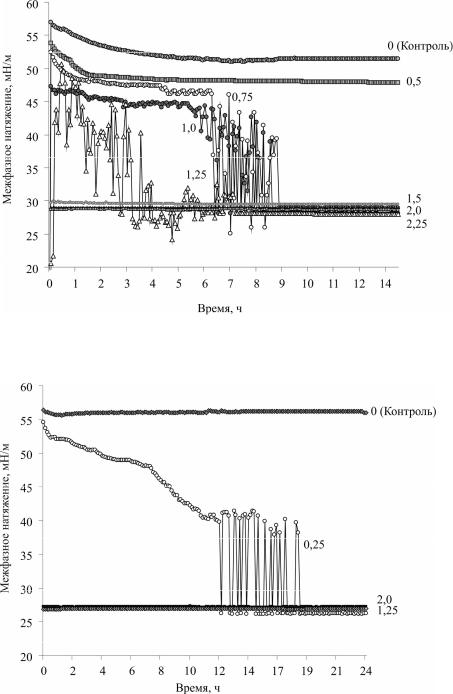
Рис. 2. Динамика межфазного натяжения суспензии клеток Rhodococcus ruber ИЭГМ 231 в системе вода-н-гексадекан. ОП600нм клеточных
суспензий: 0; 0,5; 0,75; 1,0; 1,25; 1,5; 2,0; 2,25
Рис. 3. Динамика межфазного натяжения суспензии клеток Rhodococcus ruber ИЭГМ 231 в системе вода-н-додекан. ОП600нм клеточных суспензий: 0; 0,25; 1,25; 2,0
223
Полученные результаты свидетельствуют о том, что инициация формирования биопленки в системе вода-н-додекан начинается при меньших концентрациях клеточной суспензии, что, по-видимому, обусловлено термодинамическими свойствами системы с додеканом. Вероятно, площадь раздела фаз возрастает с увеличением дисперсности системы, поэтому площадь контакта клеток с н-додеканом увеличивается и клетки родококков за более короткое время адсорбируются на доступной поверхности. Замечено, что стабильные показатели межфазного натяжения клеточной суспензии одинаковы (29–27 мН/м) независимо от природы гидрофобного субстрата.
Полученные данные о корреляционной зависимости показателей межфазного натяжения от концентрации клеток в суспензии свидетельствуют, что родококки на границе раздела фаз вода – углеводород проявляют свойства поверхностно-активных частиц [2]. Следует отметить, что бактериальные клетки, используемые в эксперименте, предварительно были отмыты от богатой питательной среды, поэтому участие Rhodococcus-биосурфактантов в данном процессе исключается. Таким образом, снижение показателей поверхностного и межфазного натяжения обусловлено участием гидрофобных взаимодействий между бактериальными клетками и углеводородом. Полученные данные могут быть использованы для регуляции роста бактериальных биопленок в жидких гетерофазных системах.
Детерминистическая биомеханическая модель временнóй зависимо-
сти поверхностного натяжения. Пусть n – концентрация клеток на межфазной границе, t – время. Будем считать, что
n n1(1 exp( t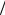 t1)) ,
t1)) ,
(n) 0 0 1(s(n)) n n1 ,
n1 ,
где n1 – предельная концентрация; t1 – «характерное время»; – межфазное (поверхностное) натяжение,
1(s) 10 ( 10 11)s,
0 s 1 – степень образования единой пленки [3–5] на межфазной границе;0 , 10 , 11 – «характерные» значения σ. Зависимость s от n определяется следующей феноменологической формулой:
s(n) 1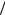 2 1
2 1 arctg (n n2 )
arctg (n n2 ) n3 ,
n3 ,
где n2 , n3 – «характерные» значения n .
Сравнивая предлагаемую теоретическую зависимость (t) с соответствующей экспериментальной зависимостью при ОП600нм = 0,75, находим t1 , 0 ,
224
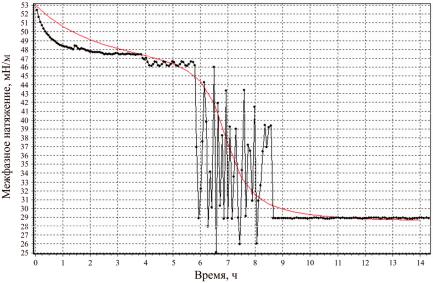
10 , 11 , n2 , n3 , обеспечивающие наилучшее совпадение без учета случайных вариацийэкспериментальнойзависимости(рис. 4).
Рис. 4. Экспериментальная зависимость σ(t) (ОП600нм = 0,75) и соответствующая теоретическая зависимость, не учитывающая случайных вариаций (плавная линия)
Вероятностная биомеханическая модель временнóй зависимости по-
верхностного натяжения. Вероятностная модель учитывает случайный характер процесса образования единой пленки бактерий. Пусть степень образования этой пленки есть случайная величина (n), равномерно распределен-
ная на отрезке s(n) (n), s(n) (n) , где
(n) k(n)min(s,1 s),
а k(n) находится из условия наилучшего (качественного) совпадения) экспериментальной и теоретической зависимостей (n) :
1,7(n 0,985), k(n) 0(n 0,985).
Сравнивая предлагаемую теперь случайную теоретическую зависимость(t) с соответствующей экспериментальной зависимостью при ОП600нм = 0,75,
находим t1 , 0 , 10 , 11 , n2 , n3 , обеспечивающие наилучшее совпадение с учетом случайных вариаций экспериментальной зависимости. Сопоставление
225
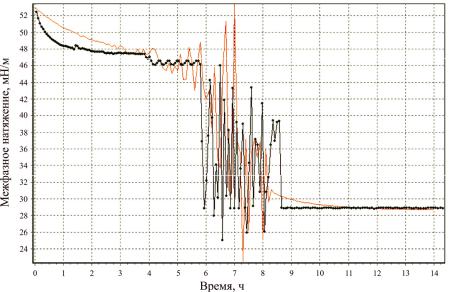
теоретической (линия без точек) и экспериментальной зависимостей представлено на рис. 5.
Рис. 5. Экспериментальная зависимость σ(t) (ОП600нм = 0,75) и соответствующая теоретическая зависимость, учитывающая случайные вариации
Сделаем следующие выводы. Экспериментально наблюдаемая зависимость межфазного натяжения в системе бактерии – вода – углеводород от времени носит сложный и нерегулярный характер. Детерминистическая модель может лишь качественно описать эту зависимость. Более точное соответствие между теорией и экспериментом может быть достигнуто только с помощью разработанной вероятностной модели. Эта модель качественно и количественно описывает основные черты рассматриваемого явления. Однако некоторые теоретические зависимости, содержащиеся в модели, являются феноменологическими, т.е. не выводятся из детального анализа механизма их возникновения. Модель допускает дальнейшее свое развитие, которое должно состоять в подробном анализе механизмов поведения бактериальных клеток на границе раздела фаз.
Список литературы
1. Ившина И.Б., Пшеничнов Р.А., Оборин А.А. Пропанокисляющие родококки / Академия наук СССР, Урал. науч. центр. – Свердловск, 1987. – 125 с.
226
2.Николаев Ю.А., Плакунов В.К. Биопленка – «город микробов» или аналог многоклеточного организма? // Микробиология. – 2007. – Т. 76, № 2. –
С. 149–163.
3.Mechanical properties of hexadecane–water interfaces with adsorbed hydrophobic bacteria / Z. Kang, A. Yeung, J.M. Foght, M.R. Gray // Colloids and Surfaces B: Biointerfaces. – 2008. – Vol. 62. – P. 273–279.
4.Hydrophobic bacteria at the hexadecane–water interface: examination of micrometre scale interfacial properties / Z. Kang, A. Yeung, J.M. Foght, M.R. Gray // Colloids and Surfaces B: Biointerfaces. – 2008. – Vol. 67. – P. 59–66.
5.Behavior of Marinobacter hydrocarbonoclasticus CP17 cells during initiation of biofilm formation at the alkane–water interface / B. Klein, P. Bouriat, P. Goulas, R. Grimaud // Biotechnology and Bioengineering. – 2010. – Vol. 105, no. 3. – P. 461–468.
Получено 1.09.2014
Феоктистова Екатерина Валерьевна – магистрант, ПНИПУ, ФПММ,
гр. БМ-13-1м, e-mail: ekaterina_feoktistova@bk.ru.
Куюкина Мария Станиславовна – доктор биологических наук, профессор, ПНИПУ, ведущий научный сотрудник Института экологии и генети-
ки микроорганизмов УрО РАН, e-mail: kuyukina@iegm.ru.
Осипенко Михаил Анатольевич – кандидат физико-математических наук, доцент, ПНИПУ, e-mail: oma@theormech.pstu.ac.ru.
227
УДК 531/524:[57+61]
Н.С. Шабрыкина, С.В. Иванишин, М.А. Гуляев
N.S. Shabrykina, S.V. Ivanishin, M.A. Gulyaev
Пермский национальный исследовательский политехнический университет
Perm National Research Polytechnic University
МОДЕЛИРОВАНИЕ ДВИЖЕНИЯ НОГИ ЧЕЛОВЕКА ПРИ ВЫПОЛНЕНИИ СПОРТИВНЫХ ДВИЖЕНИЙ
MODELING OF HUMAN BODY MOVEMENTS IN SPORT
Построена многозвенная модель ноги человека с учетом действия шести основных мышц. Модель использована для описания движения тела человека при отталкивании перед прыжком. Найдены реакции в голеностопном, коленном и тазобедренных суставах, реакция опоры и суставные моменты в зависимости от времени.
In this article a model comprising four body segments and six muscles was created. The model was used to describe body movement during the take-off in vertical jumping. Ankle, knee and hip joint reactions and torques along with ground reaction force were calculated as functions of time.
Ключевые слова: биомеханика, реакция в суставе, реакция опоры, суставной момент, фигурное катание.
Keywords: biomechanics, joint reaction force, ground reaction force, joint torque, figure skаting.
Нижние конечности выполняют опорную, рессорную и двигательную функции человеческого тела. За счет суставов, мышц и связок нижние конечности как бы амортизируют движения тела и ослабляют передачу на туловище всех толчков, сотрясений при ходьбе, беге, прыжках. При различных движения человека, как бытовых, так и спортивных (ходьба, прыжки, бег), суставы человека испытывают нагрузки, как физиологические, так и патологические. Особенно велика вероятность проявления патологических нагрузок в спорте. Для изучения нагрузок на нижние конечности человека, в том числе при занятиях спортом, используются многозвенные модели нижней конечности человека, в которых кости представляются стержнями, а суставы шарнирами [1–4].
Целью данной работы является моделирование движения нижней конечности человека при различных движениях, в частности при прыжке. В данной работе рассматриваются прыжки в фигурном катании. Наибольшие нагрузки
228
на суставы возникают при отталкивании перед прыжком и при приземлении после прыжка [2, 3]. Математическое моделирование этих фаз прыжка может помочь избежать травм и выбрать более рациональную стратегию выпрыгивания и приземления.
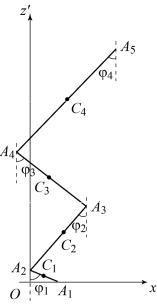
В данной работе построена четырехзвенная модель человеческого тела, которая состоит из туловища, бедра, голени, стопы (рис. 1). В модели приняты следующие допущения:
Рис. 1. Четырехзвенная модель тела человека
1.Тело человека представлено как последовательность сегментов, являющихся абсолютно твердыми стержнями.
2.Масс-инерционные характеристики сегментов соответствуют антропометрическим характеристикам частей тела человека и постоянны в течениие рассматриваемого промежутка времени.
3.Суставы представляют собой шар-
ниры.
4.Ограничения на движения суставов отсутствуют.
5.Расчеты проводятся с учетом влияния шести основных мышц ноги.
Точками С1 – С4 обозначены центры масс каждого звена, 1 4 – углы между
звеньями и вертикалью, которые определяют положение тела в пространстве. Координаты центров масс вычисляются по формулам:
|
|
i 1 |
|
|
|
xi l1 sin 1 |
( 1)k lk sin k ( 1)i ri sin i , |
|
|
|
|
k 1 |
|
|
|
|
i 1 |
|
|
|
zi |
lk cos k ri cos i , |
|
|
|
|
k 1 |
|
|
где (xi , zi ) |
– координаты центра масс i-го звена, i 1...4; |
A1C1 r1 , |
A2C2 r2 , |
|
A3C3 r3 , |
A4C4 r4 ; li – длины соответствующих звеньев, i 1...4 . |
|
||
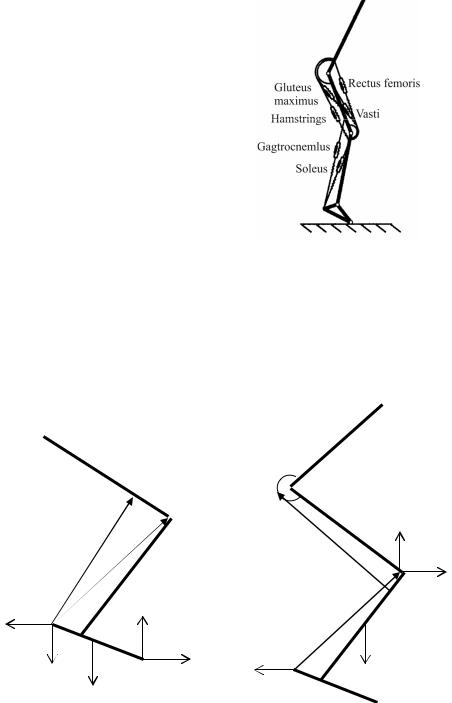
Поскольку существенное влияние на движение нижней конечности оказывают мышцы, для изучения реакций в суставах необходимо учесть действия основных мышц ноги человека. На рис. 2 изображены рассматриваемые нами мышцы.
229
Для определения реакций в суставах и суставных моментов конструкция разбивается на четыре звена. На рис. 3–6 изображены силы, действующие на каждое из звеньев. Уравнения динамики плоскопараллельного движениядлязвеньевимеютвид:
|
|
mai F , |
(1) |
|
|
Ii i M e , |
(2) |
где |
a |
– ускорения |
центров |
|
i |
|
|
масс звеньев системы, |
i 1...4; |
||
F |
– внешние силы, действую- |
||
щие на данное звено; |
Ii – мо- |
||
менты инерции звеньев относительно центра масс данного
звена; M e – моменты внешних сил, действующих на звено.
|
Fgas Fsol |
F2x |
F1x |
|
|
|
F1z |
|
F2z |
|
m1g |
Рис. 3. Силы, действующие на стопу
230
Рис. 2. схема зацепления основных мышц ноги человека: gastrocnemius – икроножная мышца (gas); gluteus maximus – большая ягодичная мышца (glu); rectus femoris – прямая мышца бедра (rec); soleus musculus – камбаловидная мышца (sol); musculus vastus medialis (vasti) – широкая мышца бедра(vas); musculus biceps femoris (hamstrings) –
двуглаваямышцабедра(ham) [5]
F3x
Fham
F3z
F2x |
Fsol |
|
m2g |
||
|
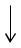 F2z
F2z
Рис. 4. Силы, действующие на голень