

Синергетика методические рекомендации
..pdfДалее следует остановиться на электрических импульсах возбуждения
– потенциалы действия распространяются по нервным и мышечным волокнам без затухания, так как в каждой точке возбудимой активной среды, до которой дошло возбуждение, заново генерируется потенциал действия. Мышечные и нервные волокна являются средами с распределенными источниками энергии метаболизма клеток. Для описания процесса распространения нервного импульса по аксону необходимо представить полный ток через мембрану m:
|
|
1d 2 |
|
I |
m |
m |
, |
|
|||
|
r dx2 |
||
|
|
||
|
|
i |
|
где ri – сопротивление аксоплазмы на единицу длины. Тогда зависимость m (x, t) описывается уравнением
1 d 2 |
|
|
|
|
|
|
d |
m |
|
|||
|
|
m |
I |
k |
I |
Na |
I |
ут |
C |
|
, |
|
|
|
|
|
|||||||||
r |
|
dx2 |
|
|
|
dt |
||||||
|
|
|
|
|
|
|
||||||
i |
|
|
|
|
|
|
|
|
|
|
|
|
где С – емкость мембраны, приходящаяся на единицу длины волокна,  – ионные токи через мембрану, Iут – ионные токи утечки, Ii –ионные токи через мембрану. Воспользовавшись уравнением Ходжкина – Хаксли, можно получить для аксона:
– ионные токи через мембрану, Iут – ионные токи утечки, Ii –ионные токи через мембрану. Воспользовавшись уравнением Ходжкина – Хаксли, можно получить для аксона:
1 d 2 |
|
d |
m |
|
p |
|
p |
|
||
|
|
m |
C |
|
gNa m |
gk m |
I ут. |
|||
r |
|
dx2 |
dt |
Na |
K |
|||||
i |
|
|
|
|
|
|
|
|
|
|
Уравнения, описывающие распространение волны возбуждения по структурам сократительного миокарда, существенно усложняются тем, что в кардиомиоците потенциал действия формируется дополнительно медленными входящими токами, сложными процессами сопряжений токов в нем.
Далее необходимо остановиться на понятии активной среды, где каждая точка есть дополнительный источник изменений диффундирующих реагентов. Затем рассмотреть вопросы самоорганизации в открытых системах с распределенными параметрами, иметь представление о диссипативной структуре и условиях ее образования.
ТЕМА 7. Биоэлектрические потенциалы
При изучении темы нужно дать краткую характеристику строения и физико-химических свойств биологических мембран и их структурных компонентов – белков и липидов. Затем следует описать подвижность структурных элементов мембран и динамику их взаимодействий, остановиться на особенностях фазовых переходов, перейти к реакциям свободнорадикального окисления липидов в мембранах, отметить роль активных форм кислорода и разобрать механизмы защиты, рассмотреть вопрос об антиоксидантах как регуляторах уровня окислительных процессов в клетках.
11
Кроме того, необходимо остановиться на модельных мембранных системах, перечислить и кратко охарактеризовать важнейшие для биофизических работ модели: монослои на границе раздела фаз, бислойные липидные мембраны, протеолипосомы; понять роль этих моделей в изучении биологических процессов, протекающих в функционирующих клетках.
Следует усвоить пути транспортировки неэлектролитов – диффузию, проникновение через поры, облегченную диффузию, понятие коэффициента распределения ионов между фазами, а также определение коэффициента проницаемости. После этого нужно специально остановиться на вопросах транспортировки ионов, разобрать понятие электрохимического потенциала, показать, что градиент электрохимического потенциала является движущей силой пассивного транспорта ионов, привести условия электрохимического равновесия и дать уравнение Нернста для равновесного потенциала. Нужно подчеркнуть, что природа энергетического барьера для переноса иона через мембрану связана с низким значением диэлектрической постоянной.
Следует рассмотреть ионное равновесие на границе раздела фаз. Отметить, что вследствие низкой и неодинаковой липофильности катионов и анионов распределение их между фазами неравномерно, и поэтому между объемами фаз возникает межфазовая разность потенциалов. Показать, что условия электронейтральности выполняются для объемов фаз, но не выполняются вблизи границы раздела. Затем следует охарактеризовать профиль потенциала в фазах вблизи раздела и профиль распределения ионов.
Следует детально рассмотреть Доннановское равновесие для двух электронейтральных фаз, показать, что возникновение потенциала связано с тем, что одна из фаз содержит заряженные частицы, не проникающие через мембрану, привести формулу для вычисления Доннановской разности потенциалов.
Далее надо по возможности подробно рассмотреть механизмы пассивного и активного транспорта ионов.
Изучая пассивный транспорт, необходимо привести уравнение зависимости скорости потока ионов от величины движущей силы. Следует подчеркнуть, что приведенное уравнение основано на термодинамических соотношениях Онзагера.
Уравнение Гольдмана описывает суммарный поток, который складывается из потоков ионов в разных направлениях. Отдельные однонаправленные потоки можно измерять с помощью изотопов. Следует получить уравнение однонаправленных потоков и показать, что поток ионов внутрь клетки зависит только от концентрации данного иона в среде, а поток наружу – только от концентрации данного иона внутри клетки. Отсюда надо вывести принцип независимости потоков и критерий Уссинга. Критерий Уссинга используется для выявления активной и пассивной компонент ионного транспорта.
Далее надо вывести уравнение мембранного потенциала из уравнения для ионных потоков. В условиях покоя суммарный ток через мембрану равен
12
нулю. Решая уравнение Гольдмана для этих потоков, получим выражение для мембранного потенциала покоя. Непосредственной причиной мембранного потенциала (потенциала покоя) является вклад неравномерного распределения ионов и различия их проницаемости в мембране:
Рк+ : РNa+ : РCl+= 1 : 0,04 : 0,45.
Следует привести данные о величине трансмембранного потенциала, который совпадает с равновесным потенциалом для ионов калия (около 70 мВ), сделать вывод о том, что отсутствие видимого вклада иона натрия в потенциал покоя объясняется работой активного механизма по удалению натрия из клетки.
После этого надо перейти к описанию механизмов активного транспорта. Необходимо изложить результаты опытов, в которых исследовалось возникновение разности потенциалов на коже лягушки, подчеркнуть, что при компенсации разности потенциалов от внешнего источника критерий Уссинга не выполняется, что указывает на наличие активного транспорта ионов натрия. Этот вывод был подтвержден прямыми опытами с использованием радиоактивных изотопов.
Специально следует остановиться на вопросе о каналах и переносчиках. Вначале надо дать общую характеристику каналов в биологических мембранах, привести их размеры, число на единицу площади, отметить, что соотношение количества натриевых каналов и калиевых каналов в возбудимой мембране равно приблизительно 10:1.
Необходимо привести описание дискретного однорядного транспорта ионов, объяснить природу энергетического барьера и модификации его профиля под влиянием разности потенциалов, далее остановиться на понятии «проводимость», которая зависит от наличия в канале свободных мест, а не только от высоты энергетического барьера, рассмотреть явление блокировки и насыщения каналов. Следует подчеркнуть, что энергетические профили каналов могут меняться под влиянием находящегося в канале иона или за счет других факторов, привести энергетический профиль натриевого канала и дать топографическую схему строения канала, разобрать механизм прохождения ионов через канал при условии дегидратации иона. Показать, как в этом случае может быть обеспечена селективность прохождения.
Надо подробно разобрать современные представления о механизме активного транспорта натрия, привести общую схему работы Na+-, K+- АТФазы, подчеркнув необходимость конформационных изменений белка для переноса ионов. Затем надо остановиться на вопросе о механизме активного транспорта кальция.
Далее надо перейти к вопросу о транспорте ионов в возбудимых мембранах, а затем рассмотреть картину возникновения потенциала действия (ПД) в нервном волокне. Указать, что ПД является натриевым, равновесным
13
потенциалом. Специально надо отметить роль ионов кальция и хлора в генерации ПД на других возбудимых объектах.
Затем следует дать общее представление о механизме распространения возбуждения в нервных волокнах. Сначала следует описать кабельные свойства нервных волокон и привести электрические параметры, характеризующие немиелиновые и миелиновые волокна. Далее изложить теорию местных токов, согласно которой возникающие между возбужденным и невозбужденным соседними участками волокна токи сами служат раздражителями и обеспечивают распространение импульса. Привести экспериментальные доказательства этой теории, показав зависимость скорости проведения от сопротивления окружающей нерв среды. Указать скорости распространения возбуждения у разных объектов – от 2–3 см/с в клетках водоросли нителла до 120 см/с в быстропроводящих нервных волокнах млекопитающих. Показать связь скорости проведения возбуждения с морфологическими и электрическими параметрами нервных волокон и отметить, что увеличение скорости проведения достигается в эволюции двумя путями – за счет увеличения диаметра аксона и за счет повышения степени миелинизации. Осветить вопрос об энергообеспечении ритмического возбуждения, указать на АТФ как основной источник энергии, привлечь данные о значении работы митохондрий, в частности электронтранспортной цепи, для энергообеспечения процесса распространения ритмического возбуждения.
На примерах биоэлектрогенеза в конечном итоге студент должен усвоить, что биоэлектрическая активность (например, потенциал действия) является результатом кооперативного (синергетического) движения многих процессов.
ТЕМА 8. Основы биологической теории информации
В результате освоения темы нужно знать, что теория информации является математической теорией с высокой степенью общности. Она основывается на теории случайных событий, для описания которых применяются понятия «вероятность» и «энтропия». В рамках самой теории вводится понятие «информация» и устанавливается ее мера – бит. Строится теория информации подобно другим теориям в математике: сначала аксиоматически определяются исходные понятия, а затем из них путем рассуждений доказывается справедливость новых положений или теорем – именно таким путем шел основоположник данной теории Клод Шеннон.
Отдельно следует остановиться на практической применимости положений и следствий, выводимых в теории, рассмотреть примеры использования теории информации в информатике, технике, психологии, биологии, физике, педагогике, лингвистике и т.д. Показать, что, как и любая математическая теория, теория информации применима для решения конкретных задач практики в той мере, в какой описываемые материальные системы или про-
14
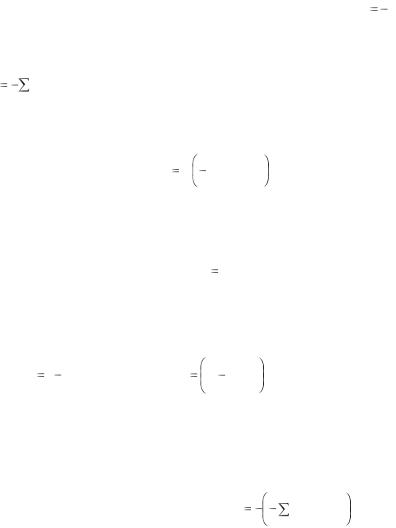
цессы удовлетворяют исходным положениям теории. Остановиться на том, что в системе, действующей без помех, вероятность (Р) равна единице, а ин-
формация (n), содержащаяся в сообщении, равна – log2 P , т.е. n |
log2 P. |
В случае, когда состояние системы определяется несколькими сообщениями, каждому из которых соответствует своя вероятность, среднее количество информации (Н), приходящееся на одно сообщение или событие, составляет
|
in |
H |
Pi log 2 Pi . В этом выражении Н называется информационной энтро- |
|
n |
пией. Если информация известна заранее (Р=1), информационная энтропия принимает минимальное значение, а в случае, если все элементы информационной системы равновероятны, энтропия принимает максимальное значение:
H max m |
1 |
log 2 |
1 |
, |
|
m |
|||
|
m |
|
||
где Нmax – максимальная энтропия в системе; m – количество элементов этой системы. Студент должен знать, что отношение энтропии информации данного состояния системы (Н) к максимально возможной энтропии для этой системы (Нmax) называется относительной энтропией, или сжатостью информации:
H |
D. |
|
|
||
Hmax |
||
|
Сопоставление энтропии (Н) с относительной энтропией (D) дает возможность судить о надежности действующей системы связи. Для количественной характеристики этой ситуации определить избыточность (R) по формуле:
R 1 |
H |
, в процентах R 1 |
H |
100. |
|
|
|
||||
Hmax |
Hmax |
||||
|
|
|
Далее рассмотреть порядок определения энтропии и избыточности информации, в случае если имеется неравномерный поток сигналов, заключается в расчетах вероятности и логарифмической функции для каждой группы одновременных сигналов. Информационная энтропия, как и термодинамическая, связана с вероятностью состояния системы и является показателем степени ее упорядоченности. Поэтому можно написать, что S = –H, или
|
in |
более полное выражение для энтропии: S |
Pi log 2 Pi . Чем менее |
|
n |
организованной и упорядоченной будет система, тем более высокое значение приобретает ее энтропия.
В заключение нужно показать, что определение энтропии системы в динамике с позиции регуляции тех или иных процессов в организме, или, точнее, всей суммы, дает возможность выяснить упорядоченность этих про-
15
цессов. Вместе с тем, чем меньше избыточность, тем менее надежным является состояние системы.
ТЕМА 9. Ритмический характер и колебательные процессы в биологии как способ организации живой системы во времени
В процессе изучения темы следует уяснить, что ритмы подразделяются по принадлежности к классу явлений (ритмы живой и неживой природы; ритмы растений, животных, человека и т.д.) и по признаку величины периода (микро-, мезо-, макроритмы). Далее перейти к классификации И.А. Тюрина, который предложил различать ритмы «экологические», или адаптивные (суточные, приливно-отливные, лунные, сезонные) и «функциональные» (все остальные ритмы, обеспечивающие осуществление программы жизни и реакции на внешние непредсказуемые воздействия).
Необходимо привести классификацию биоритмов человека по признаку принадлежности к определенному классу явлений, разработанную на основе выделенных А.Д. Слонимом уровней гомеостатических механизмов. Показать, что биологические ритмы присущи практически любому процессу.
Далее следует рассмотреть классификацию биоритмов по Халбергу, который разделил ритмические процессы на три основные группы: 1) ритмы высокой частоты с периодом до 0,5 ч; 2) ритмы средней частоты с периодом 0,5 ч – 6 дней, куда входят ультрадный ритм (период 0,5–20 ч), циркадный (период 20–28 ч), инфрадный (период 28 ч – 6 дней); 3) ритмы низкой частоты, включающие циркасептанный (недельный) ритм, циркавижинтанный (20-дневный); циркатригинтанный ритм (лунный) с периодом около 30 дней и циркадный (годичный) ритм.
Необходимо знать, что принцип обеспечения устойчивости неравновесного термодинамического состояния клеток заключается в том, что величина их неравновесного термодинамического состояния всегда непрерывно колеблется в пределах, определяемых аллостерическими модуляторами и гормонами. Поскольку каждая клетка представляет собой полноценный микроорганизм, находящийся в устойчивом неравновесном термодинамическом состоянии, образуемые этими клетками органы, системы и целостные организмы также находятся в устойчивом неравновесном термодинамическом состоянии. Причем, поскольку все биохимические процессы в этих клетках взаимосвязаны, то функционирование органов, систем и целостных организмов обеспечивается путем соответствующих совокупных синхронных интегральных колебаний неравновесного термодинамического состояния клеток.
Это находит свое выражение в периодических колебаниях физиологических параметров (функциональных сдвигов) органов, систем и целостного организма. Наглядным примером может служить последовательность сокращений и расслаблений сердечной мышцы: при сокращении сердечной мышцы у входящих в нее клетках происходят синхронные процессы расщепления АТФ, а при расслаблении – процессы синтеза АТФ. Причем при последова-
16
тельных циклах сокращения и расслабления сердечной мышцы в этих процессах статистически достоверно одновременно, интегрированно и синхронно производят соответствующие биохимические реакции огромное количество клеток, каждая из которых в составе сердечной мышцы выполняет свою роль.
При этом частота сердечных сокращений определяется термодинамическим состоянием всего организма и может колебаться в зависимости от испытываемой организмом нагрузки в достаточно больших пределах. Аналогично происходят соответствующие колебательные процессы в системе дыхания, центральной нервной системе и других. Живые организмы являются открытыми термодинамическими системами, в которых непрерывно происходят разнообразные необратимые процессы.
Поэтому условия существования и жизнедеятельности каждой клетки непрерывно меняются и соответственно меняются (перераспределяются) их роли в интегральных процессах, происходящих в органах и системах. Однако статистически, в результате действия огромного количества клеток, они в конечном итоге производят те действия, для которых предназначены в данном случае последовательные сокращения и расслабления сердечной мышцы в необходимом ритме. При восприятии импульсов света или звука периоды колебаний составляют доли секунд.
При некоторых видах труда периоды вызванных ими колебаний (в сочетании с другими колебаниями) могут составлять сутки, недели и даже месяцы. При этом, независимо ни от чего, все виды физиологических колебаний представляют собой непрерывные последовательности циклов, соответствующие энергетическим колебаниям потребления и выделения энергии. Принцип обеспечения устойчивости неравновесного термодинамического состояния живых организмов (биологических систем) как на уровне клеток, так и на уровне органов, систем и целостных организмов заключается в непрерывных чередованиях потребления и выделения энергии посредством управляемых на соответствующих уровнях циклов синтеза и расщепления АТФ.
Необходимо ознакомиться с современными представлениями о природе биологических ритмов, о факторах-синхронизаторах биологических ритмов, о роли ритмов в качестве механизма адаптации в жизнедеятельности организмов от простейших до человека, о механизмах регуляции биологических ритмов.
Следует уяснить, что такое временная организация биологических систем, какие методы существуют для ее изучения, иметь представление о ритмической структуре среды обитания, современном состоянии проблемы биологических часов и регуляции биологических ритмов.
Показать возможные причины универсальности колебательных процессов в природе. Осветить вопрос автоколебания и автоволны. Перейти к понятиям, которыми можно описать биологический ритм. Знать типы спектров и их зависимость от сложности сигнала. Далее остановиться на понятиях «синхронизация», «резонанс» и разграничение их типов.
17
ТЕМА 10. Начало физиологической синергетики
При изучении темы особое внимание нужно обратить на то, что в динамике напряжения кислорода, импульсной электрической активности и ионов йода значительную роль для осуществления физиологических функций играют ритмические и аритмические их изменения, названные флуктуациями. Между различными по энергоемкости, скорости и принадлежности к физиологическим системам регулирования микрофизиологическими показателями энергопродукции, энергопотребления и агрегатного состояния митохондрий происходит амплитудно-частотная синхронизация, которая является одним из физиологических механизмов адаптации к гипоксии или иному действующему на организм фактору природы. Передача синхронизированных сигналов действия может осуществляться акустическим методом – например, амплитудно-частотные звуковые сигналы от пульса, т.к. именно пульс обладает уникальной информацией о синхронной или асинхронной работе клеток организма, поскольку кровь омывает все органы, а пульс несет информацию о них.
ТЕМА 11. Квантово-волновые механизмы синергетических процессов
В процессе освоения темы следует уяснить основы организации процессов синхронизации и десинхронизации, основные категории квантововолновой теории, образование радикалов.
Следует составить представление о строении атомов и молекул и их энергетических уровнях. Вспомнить, что химические свойства атомов определяются строением их наружного электронного слоя. Часть электронов, находящихся во внешнем электронном слое атомов, являются спаренными, а часть – неспаренными. Спаренные электроны находятся на одной орбите и имеют антипараллельные спины. Неспаренные – это одиночные электроны на орбитах.
При образовании молекул химическая связь возникает в основном благодаря взаимодействию неспаренных электронов двух атомов – первые признаки синхронизации и кооперации на уровне атомов. В основном все электроны в молекуле спарены, и их суммарный спиновой эффект равен нулю. Такое состояние электронных оболочек называется синглетным. Если молекула имеет один неспаренный электрон, то такое соединение называется монорадикалом, если два неспаренных электрона – то бирадикалом.
Важнейшим для организма веществом, основное состояние которого бирадикально, является кислород – его молекула обладает двумя неспаренными электронами. То обстоятельство, что молекула кислорода содержит два неспаренных электрона, одновременно принадлежащих двум ее атомам и имеющих одинаково направленные спины, придает молекулярному кислороду парамагнитные свойства. Свободные радикалы могут быть нейтральными и заряженными.
18
Кроме электронной энергии, молекулы обладают колебательной и вращательной энергией, относящейся к тепловой энергии. Колебательная энергия молекул обусловлена колебаниями отдельных атомов или групп атомов внутри молекулы. Вращательная энергия молекул обусловлена вращением молекул вокруг каких-либо осей. Колебательная и вращательная энергия молекул, как и электронная энергия, имеет квантовый характер, т.е. изменяется дискретно. Квантово-волновые свойства синглетов, радикалов, парамагнитных молекул кислорода, вращения и колебания молекул создают синхронозы и десинхронозы (синергетика) – основа возникновения значительных флуктуаций в биосистемах, которые могут привести к различным патологиям.
ТЕМА 12. Применение методов синергетики
При изучении темы следует уяснить, что лазер это оптический квантовый генератор. В межзвездном пространстве обнаружена природная лазерная генерация. Необходимо рассмотреть работу рубинового лазера. При малой мощности накачки лазер работает как обычный источник света. Начиная с некоторого порогового значения мощности накачки, все атомы согласованно испускают свет в одной фазе, т.е. возникает кооперативное поведение атомов и излучения. В системе происходит самоорганизация. Некогерентный (неупорядоченный) свет накачки изменяет свои свойства, трансформировавшись в организованный свет лазерного излучения. Он становится когерентным, усиленным в направлении испускания, более узким в пространственном и спектральном отношении.
Далее необходимо рассмотреть систему, представляющую собой слой жидкости (воды) между двумя горизонтальными пластинами. При постоянной температуре жидкость находится в однородном равновесном состоянии. Если верхняя пластина остается холодной, а нижняя подогревается, то в пределах некоторой разности температур может осуществляться стационарный процесс теплопроводности, не меняющий однородную структуру жидкости. Однако при разности температур состояние меняется. Нижние теплые «капли» жидкости под выталкивающим действием архимедовой силы, преодолевая вязкость, начнут движение вверх. Верхние холодные «капли» под действием силы тяжести будут опускаться вниз. В результате возникают восходящие и нисходящие конвекционные потоки, жидкость закручивается и структурируется в виде небольших ячеек, называемых ячейками Бенара. Таким образом, система (жидкость) вместо однородной структуры сама формирует новую динамическую сложную, упорядоченную во времени и пространстве структуру, т.е. она самоорганизуется.
Затем следует привести примеры биологических систем, функционирующих вдали от равновесия, в которых просматривается связь между физи- ко-химической организацией и биологической упорядоченностью. Необходимо знать, что организм как целое непрерывно получает потоки энергии
19
(например, в виде солнечной энергии, используемой растениями при фотосинтезе) и вещества (в виде питательных веществ) и преобразует их в разные отходы, выделяемые во внешнюю среду. Особое внимание следует уделить клеточному уровню, где также наблюдаются сильные неоднородности (биоэлектрогенез). Неравновесность живых систем поддерживается за счет работы активного транспорта и биоэлектрических реакций типа гликолиза и дыхания. Далее показать, что эмбриональное развитие представляет собой последовательность событий, ведущих от отдельной клетки – оплодотворенного яйца – к целому организму, что является синергетической моделью.
ТЕМА 13. Синергетика работы головного мозга
При изучении темы необходимо показать, что мозг человека функционирует вблизи критического состояния. Чрезвычайная чувствительность мозга к малейшим изменениям, как внешних стимулов, так и внутренних психических процессов указывает на то, что мозг как сложная система функционирует вблизи бифуркационного состояния. Такие явления, как сознание и подсознание, а также творчество, нейропластичность, синхронизация как механизм интегративной функции мозга, восприятие неоднозначных образов связаны с феноменологией критических явлений. Функция распознавания – фундаментальное свойство живой природы. Примером может служить распознавание иммунной системой инородных объектов. Наиболее распространенная в синергетике модель распознавания образов, предложенная американским физиком Джоном Хопфилдом, обладает важным свойством мультистабильности, что делает эту модель согласованной со сформулированным выше принципом критического состояния, вблизи которого функционирует мозг.
Нейронная сеть способна по неполным данным восстановить полный ключевой образ, хранящийся в памяти. Условный рефлекс, способность прогнозирования будущих событий также можно описать как процесс восстановления полного образа по его фрагменту. Электроэнцефалограммы (ЭЭГ) животных и человека, а также магнитные поля, генерируемые в межнейронных тканях мозга, представляют собой детерминированные хаотические процессы с небольшим числом степеней свободы, что указывает на высокую степень самоорганизации соответствующих процессов.
Традиционные исследования рассматривают функции мозга как строго локализованные, осуществляющиеся на уровне индивидуальных нейронов, в то время как в синергетическом подходе все основные функции мозга (память, распознавание образов и т.д.) считаются распределенными среди огромного числа взаимосвязанных нейронов. Следует рассмотреть еще одно принципиальное отличие – в концепции обработки информации мозгом. Традиционные науки о мозге отдают предпочтение концепции последовательной обработки информации, в то время как синергетика мозга базируется на идее параллельной обработки.
20