

Глава 2. Структура и функции хлоропластов.
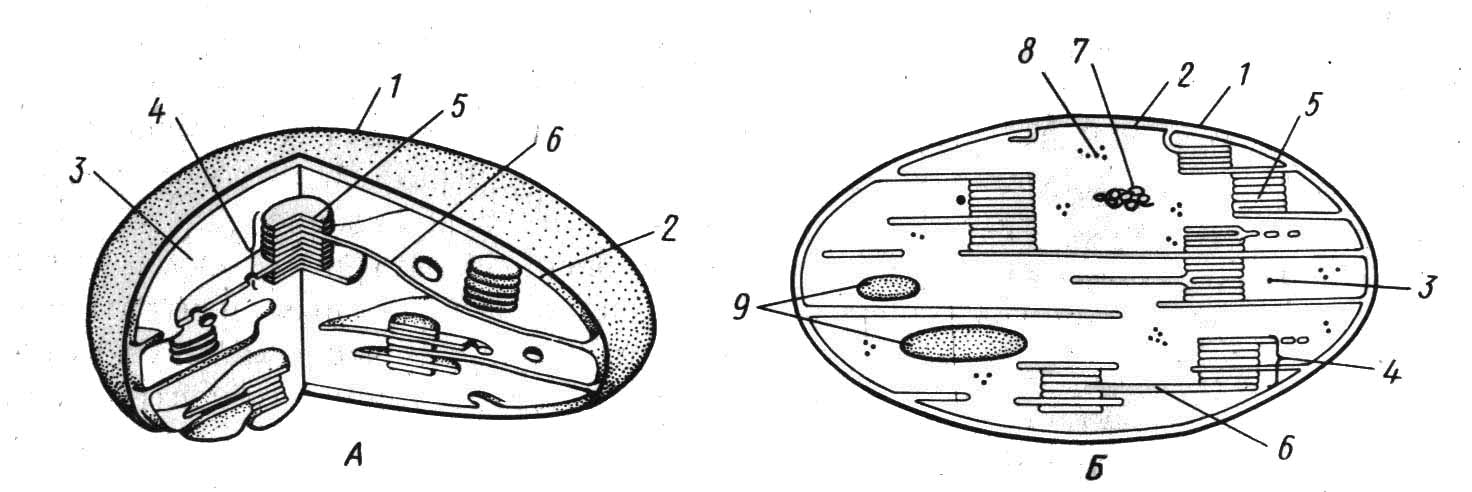
Хлоропласты – это тельца овальной формы шириной 2-4 мкм и длиной 4-10 мкм. Они имеют две мембраны толщиной по 7 нм с межмембранным пространством шириной около 30 нм. Как и у митохондрий, наружная и внутренняя мембраны хлоропласта отличаются проницаемостью и другими физико-химическими свойствами. Внутренняя мембрана хлоропластов образует протяженные складки – ламеллы. На ламеллах располагаются плоские мембранные цистерны дисковидной формы – тилакоиды, имеющие полость шириной 25 нм. Они собраны в комплексы наподобие столбика монет – граны. Тилакоиды уложены в гране таким образом, что между соседними мембранами остается пространство шириной 2 нм. Число тилакоидов в гране может достигать несколько десятков.
 Внутри
хлоропласта между мембранными структурами
содержится мелкодисперсное вещество,
формирующее матрикс, или строму. У
некоторых хлоропластов и других пластид
в строме обнаруживаются включения –
пластоглобулы, крахмальные зерна
и кристаллы белка.
Внутри
хлоропласта между мембранными структурами
содержится мелкодисперсное вещество,
формирующее матрикс, или строму. У
некоторых хлоропластов и других пластид
в строме обнаруживаются включения –
пластоглобулы, крахмальные зерна
и кристаллы белка.
1.Наружняя мембрана, 2-внутренняя мембрана, 3-строма, 4-грана, 5-тилакоиды граны, 6-тилакоиды стромы, 7-нить днк, 8-рибосомы, 9-крахмальные зерна.
Рисунок 2.1. – Хлоропласт (А-трехмерная модель, Б-срез)
Фотосинтез – основная и главная функция хлоропластов. В хлоропластах осуществляется фотосинтез, в результате которого из углекислого газа и воды с использованием энергии света образуется органическое вещество и выделяется кислород. Процесс фотосинтеза подразделяется на световую и темновую фазы.
Световые реакции. Преобразование
энергии света в энергию химических
связей начинается в реакционных центрах,
входящих в состав мембран тилакоидов.
В составе реакционных центров
обнаруживаются разнообразные сочетания
пигментов: хлорофиллы а и b, каротиноиды
и другие. Кроме указанных пигментов
в мембранах обнаруживаются разнообразные
вещества – переносчики электронов и
протонов. Основные сочетания пигментов
и переносчиков называются 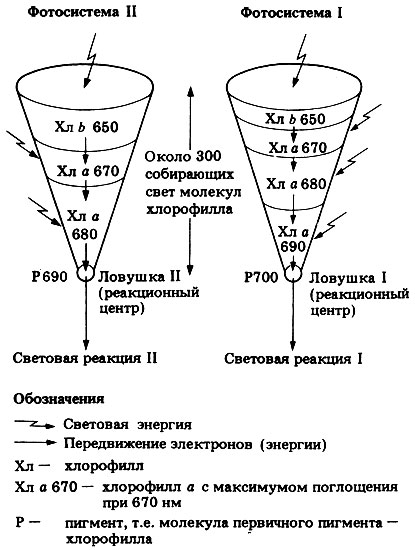 фотосистемы:
фотосистема I и фотосистема II.
фотосистемы:
фотосистема I и фотосистема II.
Рисунок 2.2. – Схематическое представление об энергетических ловушках в фотосистемах I и II.
Универсальным способом образования АТФ является механизм нециклического фотофосфорилирования. Энергия света, поглощенная пигментами, преобразуется в энергию электронов. Свободные электроны образуются при фотолизе (фотоокислении) воды – расщеплении молекулы Н2О с затратой световой энергии. При фотолизе воды выделяется молекулярный кислород. Энергия электронов используется для создания протонных резервуаров внутри тилакоидов и формирования электрохимических потенциалов на мембранах тилакоидов. В свою очередь, энергия электрохимического потенциала используется для синтеза АТФ. Электроны, потерявшие энергию, используются для восстановления НАДФ. В действительности световые реакции протекают более сложно.
Фотосистема II поглощает высокоэнергетические кванты света. Электроны хлорофилла переходят в возбужденное состояние, а затем молекула хлорофилла теряет один возбужденный электрон с избытком энергии. Окисленный хлорофилл отщепляет один электрон от молекулы воды. Вода разлагается на протон Н+ и свободный радикал НО. Два радикала НО- объединяются в молекулу Н2О2, которая разлагается каталазой на Н2О и О2.
Процесс расщепления воды под воздействием света называется фотолиз. При фотолизе выделяется молекулярный кислород как побочный продукт световых реакций фотосинтеза:
4 Н2О → 4 Н+ + 4 НО· + 4 ē; 4 НО· → 2 Н2О2 → 2 Н2О + О2↑
Высокоэнергетические электроны от молекул хлорофилла присоединяются к хинонам, образуя восстановленные хиноны (KoQ 2–). Восстановленные хиноны диффундируют на внешнюю сторону мембраны тилакоида (к строме). Здесь к хинонам присоединяются протоны, которые всегда присутствуют в водных растворах вследствие электролитической диссоциации воды. Хиноны вместе с протонами диффундируют на внутреннюю сторону мембраны (к матриксу тилакоида). Под воздействием цитохромов b протоны отщепляются от хинонов и переходят в матрикс тилакоида. Затем хиноны вновь диффундируют к строме, где вновь присоединяют протоны. Таким образом, строма служит источником протонов, а матрикс тилакоидов – протонным резервуаром. Электроны, частично израсходовавшие энергию на перенос протонов, отщепляются от хинонов и поступают на промежуточный переносчик – цитохром f.
Фотосистема I поглощает низкоэнергетические кванты света. Электроны хлорофилла фотосистемы I переходят в возбужденное состояние, а затем молекула хлорофилла теряет один возбужденный электрон. Потерю электронов молекулы хлорофилла восполняют, забирая электроны от цитохромов f. Электроны от фотосистемы I через промежуточные мембранные переносчики (ферредоксин и другие) используются для восстановления немембранного переносчика электронов и протонов НАДФ:
НАДФ+ + 2 ē + 2 Н+ → НАДФ·Н+Н+.
Избыток протонов из матрикса переходит через канал АТФазы в строму. Энергия электрохимического потенциала используется для фотофосфорилирования – синтеза АТФ из АДФ и неорганического фосфата. В итоге энергия света расходуется на синтез АТФ и на восстановление НАДФ.
Темновые реакции. АТФ и НАДФ·Н+Н+, образовавшиеся в ходе световых реакций, используются для восстановления СО2 и образования глюкозы. Образовавшаяся глюкоза превращается в первичный крахмал. Первичный крахмал в дальнейшем гидролизуется с образованием глюкозы. Эта глюкоза транспортируется за пределы хлоропласта: в остальные клетки и органы растения. Здесь она превращается во вторичный крахмал, используется для дыхания и для биосинтеза кислот, аминокислот и других веществ. Суммарное уравнение фотосинтеза записывается следующим образом:
6 СО2 + 6 Н2О + световая энергия → С6Н12О6 + 6 О2 + тепло
Существует несколько механизмов темновых реакций. Универсальным способом фиксации СО2 является цикл Кальвина. Пятиуглеродный сахар рибулозодифосфат с помощью РДФ-карбоксилазы присоединяет одну молекулу СО2. Образуется неустойчивое шестиуглеродное соединение, которое разлагается на две молекулы фосфоглицериновой кислоты (ФГК). С помощью АТФ и НАДФ·Н+Н+ каждая молекула ФГК восстанавливается до фосфоглицеринового альдегида (ФГА). Одна шестая часть молекул ФГА в ходе реакций изомеризации и димеризации образуют фруктозу, которая превращается в глюкозу. Большая часть ФГА (5/6) используется на образование рибулозодифосфата.