

Блохин Г.И., Александров В.А. Зоология
.pdfРис. 41. Схематическое изображение строения кораллового полипа Alcyonium:
/ — щупальце; 2—ротовое отверстие; 3— глотка; 4 — перегородки кишечной полости — септы; 5 — мезентериальные нити; 6 — яйца
У колониальных полипов мощный скелет чаще всего представлен углекислыми солями, реже — рогоподобным веществом. Скелет может быть наружным или внутренним.
Коралловые полипы размножаются бесполым и половым путем. Ведущие одиночный образ жизни актинии иногда размножаются делением, у колониальных видов наблюдается почкование. Половые железы формируются в перегород-
ках между энтодермой и мезоглеей. Спермин выходят через ротовое отнерстие наружу и через рот же проникают в гастральную полость женской особи, где и происходит оплодотворение. У некоторых форм опнодотворение наружное. Развитие происходит с метаморфозом: из зиготы развивается плавающая личинка — планула, которая прикрепляется к субстрату и дает начало новому полипу
Актинии — одиночные шестилучевые яркоокрашенные полипы, лишенные скелета (рис. 42). Они могут медленно передвигаться с помощью мускулистой подошвы. Актинии очень чувствительны к раздражениям, сильно сокращаются, превращаясь в небольшой комок. Это хищники, питающиеся ракообразными, моллюсками и другими крупными животными, которых они захватывают щупальцами, парализуя стрекательными нитями.
Некоторые актинии живут в симбиозе с раками-отшельниками, поселяясь на их раковинах. Рак служит для актиний средством передвижения, а актинии пассивно защищают рака от хищников.
В тропиках распространены рифообразующие мадрепоровые шес-
Рис. 42. Актиния на раковине, занятой ра> ком-отшельником
71
тилучевые кораллы, характеризующиеся крупными размерами — более 4 м высотой. Это требовательные к температуре воды и ее солености (около 3,5 % солей) животные. Очень чувствительны коралловые полипы к загрязнению воды. Коралловые рифы служат местом обитания многих морских организмов. Животные и растения в коралловых рифах образуют своеобразное сообщество (биоценоз) рифа. В клетках энтодермы коралловых полипов живут симбиотические одноклеточные водоросли — зооксантеллы. Кораллы снабжают водоросли диоксидом углерода и предоставляют им укрьггие, а от зооксантелл кораллы получают кислород и продукты фотосинтеза, которые поступают непосредственно из клеток водорослей. Умирающие водоросли перевариваются в цитоплазме полипов. Без такого сложного симбиоза коралловые полипы погибают.
Рифы условно делят на три типа: береговые, барьерные и атоллы. Береговые расположены непосредственно по берегам островов или материков, барьерные рифы располагаются параллельно береговой линии на некотором расстоянии. Атоллы — это кольцеобразные возвышающиеся над океаном коралловые острова с озерцом внутри.
Значение кишечнополостных в Мировом океане трудно переоценить: с их помощью осуществляется круговорот кальция в биосфере, они очищают морскую воду от органической взвеси, являются звеньями в пищевых цепях и т. п. Кишечнополостные служат объектом промысла: медузы (Япония, Китай), кораллы для украшений, коллекций и ювелирных изделий, медицинских препаратов.
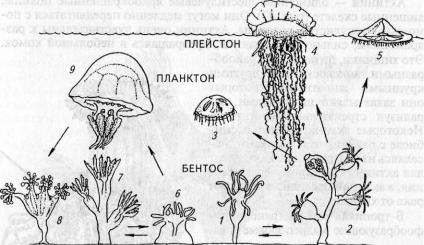
Рис. 43. Экологическая радиация кишечнополостных:
1 — гидроидный полип; 2— морской колониальный полип; 3 — гидромедуза; 4 — сифонофора; 5 — плавающий полип; 6— одиночный коралловый полип; 7— колониальный коралловый полип; 8 — сидячая медуза; 9— сцифоидная медуза
72
ФИЛОГЕНИЯ КИШЕЧНОПОЛОСТНЫХ
Кишечнополостные — древняя группа примитивных животных. Считается, что далекими предками кишечнополостных были двухслойные плавающие многоклеточные животные, похожие на планулу. Видимо, первыми были одиночные полипы. От полипов без перегородок развивались различные группы гидроидных. Коралловые полипы в процессе эволюции дали широкий спектр полипоидных форм: одиночных и колониальных, со скелетом и без него, но при этом сохранили древний признак развития без метагенеза. Основные пути морфо-экологической эволюции отражены на схеме радиации жизненных форм (рис. 43).
ТИП ГРЕБНЕВИКИ (Ctenophora)
Гребневики — это морские животные, характеризующиеся радиальной симметрией; они ведут одиночный свободноплавающий образ жизни, реже встречаются ползающие или сидячие формы. Известно около 120 видов гребневиков, заселяющих все моря. Питаются эти животные обычно планктоном.
Форма прозрачного и нежного тела мешковидная или грушевидная (рис. 44). Вдоль тела тянутся восемь рядов тонких и прозрачных гребных пластинок, образованных сросшимися ресничками. Пластинки
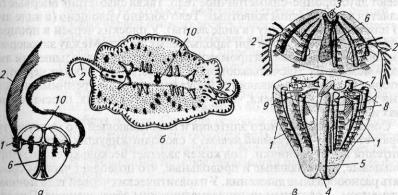
Рис. 44. Гребневики:
и — плавающий гребневик Bolinopsis; б — ползающий гребневик Coeloplana; в — схема строения гребневика; 1 — ряды гребных пластинок; 2— щупальца; 3 — орган равновесия; •/ — рот; 5—глотка; б—кишечная полость; 7—кишечные каналы, отходящие от желудка; Я — продольные кишечные каналы; 9— влагалище щупальца; 10— органы чувств
73
расщеплены наподобие гребешка и бьют по воде в одном направлении, что позволяет гребневикам двигаться только в одном направлении — ротовым концом тела вперед. Многие гребневики имеют пару щупалец с расположенными на них особыми клетками, вырабатывающими липкий секрет. С помощью этих щупалец гребневики захватывают разных животных, которыми питаются.
Рот, расположенный на оральном конце тела, ведет в глотку, которая переходит в короткий пищевод и затем в кишечную полость. Кишечная полость имеет разветвленные каналы и слепые отростки. Это двухслойные животные: снаружи тело покрыто эктодермой, а кишечная полость и ее каналы выстланы энтодермальным эпителием. Между экто- и энтодермой лежит студенистая прозрачная мезоглея. Наличие в мезоглее многочисленных клеточных элементов и отсутствие стрекательных клеток отличает гребневиков от кишечнополостных.
Гребневики — гермафродиты; половые железы расположены в стенках пищеварительных каналов. Половые клетки выходят в просвет кишечных каналов и оттуда через рот наружу. Оплодотворение наружное. В развивающемся эмбрионе происходит образование зачаточного третьего зародышевого листка — мезодермы. Развитие без метаморфоза.
ТИП ПЛОСКИЕ ЧЕРВИ (Plathelminthes)
Общая характеристика. Большинство представителей этого типа имеют двусторонне-симметричное тело. Такая симметрия впервые появляется у этой группы животных. Тело обычно уплощено (в виде листа) или вытянуто в длину (в виде ленты). У плоских червей в процессе онтогенеза формируются три зародышевых листка. Между эктодермой и энтодермой развивается промежуточный (третий) зародышевый лис- ток—мезодерма. Насчитывают около 15 тыс. видов плоских червей, из которых большинство являются наружными или внутренними паразитами животных и человека, часть червей живут в морях, пресных водоемах и почве.
Совокупность кожного эпителия и подстилающей его мускулатуры образует кожно-мускульный мешок. У свободноживущих форм кожный эпителий имеет реснички. Под кожей залегает несколько слоев мышц: кольцевые, диагональные и продольные, что позволяет червям совершать разнообразные движения. У паразитических червей тело покрыто тегументом — сложным цитоплазматическим образованием. Плоские черви не имеют полости тела (бесполостные), поскольку все пространство между внутренними органами и стенкой тела заполнено рыхло расположенными клетками мезодермального происхождения — паренхимой (паренхиматозные черви); в промежутках между клетками паренхимы циркулирует межтканевая жидкость. Паренхима выполняет опорные функции, в ней накапливаются резервные питательные вещества, она участвует в процессах обмена веществ.
74
Пищеварительный канал примитивен, обычно разветвлен и представлен двумя отделами: эктодермальной глоткой (передняя кишка) и энтодермальной средней кишкой, которая заканчивается слепо. Задней кишки и анального отверстия нет. У части паразитических форм кишечник отсутствует.
Нервная система представлена парным мозговым ганглием и отходящими от него несколькими парами нервных стволов, идущих назад и соединенных между собой кольцевыми перемычками — комиссурами. Таким образом, у плоских червей формируется центральный аппарат 11 ервной системы. Органы чувств наиболее развиты у свободноживущих видов: имеются глазки, органы равновесия — статоцисты и многочисленные сенсиллы (осязательные клетки и органы химического чувства).
Кровеносная и дыхательная системы отсутствуют. Свободноживущие плоские черви дышат через кожу; для эндопаразитических форм характерно анаэробное дыхание.
У плоских червей появляются органы выделения, построенные по типу протонефридиев в виде системы разветвленных канальцев, оканчивающихся в паренхиме звездчатыми клетками с пучком ресничек внутри. Реснички способствуют откачке конечных продуктов обмена из паренхимы в один или два магистральных канала и затем через специальные выделительные отверстия (экскреторные поры) выводу этих продуктов наружу.
Плоские черви в большинстве своем гермафродиты. Половая система устроена сложно и обеспечивает внутреннее оплодотворение и высокую плодовитость. Развитие может быть прямым или с метаморфозом. Эндопаразитам присущи сложные жизненные циклы с чередованием обоеполого и нескольких партеногенетических поколений.
К типу плоских червей относят десять классов, из которых шесть — исключительно паразитические. Наиболее многочисленными являют- ся четыре класса: Ресничные черви (Turbellaria), Дигенетические сосальщики (Trematoda), Моногенетические сосальщики (Monogenea) и Ленточные черви, или Цестоды (Cestoda).
Считается, что плоские черви произошли от древних кишечнополо- с гпых, которые перешли к передвижению по дну, где они могли вести хищнический образ жизни. Сначала основой для движения служили I >сспички, но постепенно главенствующая роль перешла к мускулатуре гена. Активный образ жизни позволил турбелляриям усложнить систе- MI.I органов. От турбеллярий позднее произошли паразитические фор- мы плоских червей.
КЛАСС РЕСНИЧНЫЕ ЧЕРВИ (Turbellaria)
К классу ресничных червей относится большая группа (около t. 'i тыс. видов) свободноживущих в воде или в почве плоских червей, гсио которых не расчленено и покрыто мерцательным (ресничным) мштелием. Все турбеллярии — хищники. На переднем конце тела рес- ничных червей имеется несколько примитивных глазков. У болыпин-
75
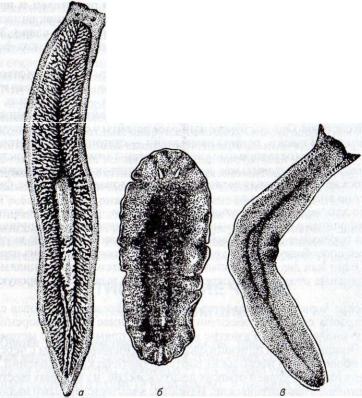
Рис. 45. Виды ресничных червей:
а — молочная планария Dendrocoelum lacteum\ 6 — морская турбеллярия Leptoplana; в — планария многоглазка Polycoelis cornuta
ства представителей рот расположен посередине тела на его брюшной стороне.
Строение и жизненные отправления. Длина тела может колебаться от долей миллиметра до 35 см. Форма тела уплощена и чрезвычайно разнообразна. Морские турбеллярии ярко окрашены (рис. 45).
Покровы представлены ресничным однослойным эпителием. Реснички способствуют передвижению мелких червей в воде, а более крупные представители ползают, вытягивая, сокращая и изгибая тело. В кожном эпителии располагаются особые палочковидные образования — рабдиты, выполняющие защитные функции: выбрасываясь наружу, они окутывают врага рыхлой клейкой оболочкой. В покровах ресничных червей много железистых клеток, одни из которых выделяют слизь, а другие — ядовитые вещества. Так, молочная планария на-
76
крывает жертву своим телом и убивает ее ядом, который вырабатывают ядовитые железы, находящиеся на брюшной стороне тела червя.
Нервная система у разных представителей различна по своей сложности. У примитивных форм она диффузного типа. Есть виды, у которых вдоль тела идет несколько нервных тяжей. У более сложно организованных имеются ганглии с продольными нервными тяжами. Органы чувств представлены примитивными глазками, статоцистами и осязательными клетками.
Органы пищеварения. У большинства видов на брюшной стороне в средней ее части расположен рот, ведущий в глотку, которая может выпячиваться наружу, присасываться к жертве и высасывать ее содержимое. Кишечник чаще всего ветвится на две и более ветвей, заканчивающихся слепо. Пища переваривается в полости кишечника и с помощью фагоцитоза (внутриклеточно) в клетках эпителия кишечной полости. Непереваренные остатки пищи выбрасываются наружу через рот. У некоторых ресничных червей кишечник отсутствует, а пища переваривается в пищеварительных вакуолях клеток, расположенных в специально обособленной пищеварительной паренхиме.
Газообмен осуществляется путем диффузии кислорода из воды через покровы внутрь тела, а диоксида улерода — наружу.
Выделительная система протонефридиального типа (рис. 46) как отдельная система органов впервые появляется у ресничных червей. У многих морских червей органов выделения нет: продукты обмена удаляются из тела через покровы и стенки кишечника.
Органы размножения устроены сложно. Большая часть ресничных червей гермафродиты. Мужская половая система представлена множе-
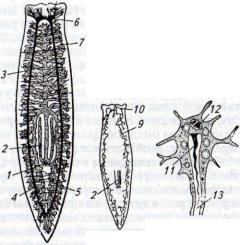
Рис. 46. Нервная система, органы пищеварения и выделения планарии:
а — пищеварительная и нервная системы; б — расположение главных канальцев выделительной системы; в — одна из концевых клеток протонефридиальной системы; / — рот; 2 — глотка; 3 — передняя ветвь кишечника; 4,5 — шдние ветви кишечника; 6 — головной нервный узел; 7—боковой нервный ствол; 8— глазок; 9— канальцы выделительной системы; К) — выделительная пора; 11 — «мерцательное пламя»; 12—ядро клетки; 13— внутриклеточный каналец
ством мелких семенников, разбросанных в паренхиме. От семенников отходят семявыносящие канальцы, которые, сливаясь, образуют два семяпровода. Семяпроводы формируют непарный семяизвергательный канал, пронизывающий совокупительный орган, расположенный в половой клоаке. Сюда же впадают и женские половые протоки.
Женская половая система представлена одним или множеством яичников. От яичников отходят два яйцевода, принимающие протоки желточников и сливающиеся в один канал — влагалище. Влагалище открывается в половую клоаку. Оплодотворенные яйцеклетки окружаются желточными клетками и вместе с ними покрываются общей скорлупой.
Благодаря разным срокам созревания половых продуктов самооплодотворения у этих червей не происходит. Оплодотворение внутреннее. Оплодотворенные яйца выводятся наружу либо через разрывы стенок тела, либо через рот, либо через специальные выводные протоки. У пресноводных форм развитие прямое. У морских видов развитие с превращением: из яйца развивается планктонная личинка, плавающая с помощью ресничек в толще воды.
Интересна способность ресничных червей к регенерации: при расчленении одного червя на сотни частей из каждой части может восстановиться новая особь.
В морях и океанах обитают мелкие ресничные черви из отрядов Бескишечные, Макростомиды, Многоветвистокишечные и др. Среди пресноводных представителей отечественной фауны можно отметить мо- лочно-белую планарию, многочисленные трехветвистокишечные турбеллярии населяют озеро Байкал. Многие виды ресничных червей служат кормом для рыб.
КЛАСС СОСАЛЬЩИКИ (Trematoda)
Общая характеристика. Известно более 4 тыс. видов сосальщиков, многие из которых являются широко распространенными и опасными эндопаразитами животных и человека. Тело сосальщиков не расчленено и имеет листовидную форму. У большинства есть присоски для прикрепления к телу хозяина. Кишечник двуветвистый, заканчивается слепо. Большинство гермафродиты. Развитие происходит со сменой хозяев. Взрослые формы живут только у позвоночных животных, поражая в основном органы пищеварения, но есть виды, обитающие в легких, кровеносной системе и в других органах. В связи со сменой хозяев в жизненном цикле сосальщиков они получили еще одно название — Digenea, т. е. двойственное развитие.
Строение и жизненные отправления. Строение сосальщиков во многом сходно со строением ресничных червей. На теле обычно имеются две присоски (отсюда и другое название сосальщиков — двуустки): одна окружает рот, а другая расположена на брюшной стороне тела (рис. 47). Иногда брюшная присоска может отсутствовать.
Покровы представлены тегументом, снаружи у некоторых форм усеянным шипиками, способствующими фиксации паразита в теле хозяина.
78
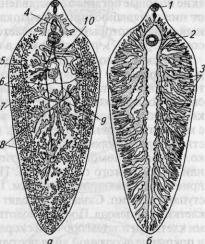
Рис. 47. Строение половой (л) и пищеварительной (б) систем печеночного сосальщика фасциолы:
/ — ротовая присоска; 2 — брюшная присоска; 3 — разветвленный кишечник (левая и правая ветви); 4 — копулятивный орган; 5 —яичник; 6— желточники; 7 — желточные протоки; 8 — семенники; 9— семяпроводы; 10— матка
Мускулатура представлена слоями мышечных волокон, которые вместе с тегументом образуют кожно-мускульный мешок. Двигаются трематоды медленно.
Нервная система слагается из парного головного ганглия и отходящих от него парных нервных тяжей. От ганглия и тяжей идут ответвления ко всем
органам. У взрослых червей органы зрения отсутствуют. В покровах размещены осязательные и другие нервные окончания.
Органы пищеварения начинаются ротовым отверстием, ведущим в глотку, которая может совершать сосательные движения. За глоткой лежит пищевод, который ветвится на две ветви кишечника. Иногда ветвистый кишечник имеет дополнительные боковые отростки, облегчающие распределение продуктов пищеварения в паренхиме червя. Анальное отверстие отсутствует
Пищеварение в основном происходит в полости кишечника (внеклеточное). Наряду с кишечным пищеварением наблюдается всасывание растворенных органических веществ через покровы тела. Непереваренные остатки пищи выбрасываются через ротовое отверстие. Продукты пищеварения транспортируются в теле сосальщиков с помощью межтканевой жидкости и клеток паренхимы за счет сокращения мускулатуры тела.
Органы дыхания отсутствуют. У эндопаразитических форм дыхание анаэробное, процессы диссимиляции происходят по типу брожения. Но есть и исключения: паразиты, живущие в легких, могут дышать через покровы своего тела.
Органы выделения протонефридиального типа. От выделительных звездчатых клеток, покрытых ресничками, и разбросанных в паренхиме, отходят канальцы, которые сливаются в более крупные выделительные каналы. Вся выделительная система каналов открывается в мочевой пузырь, а из него конечные продукты обмена веществ выбрасываются через выделительное отверстие на заднем конце тела.
Органы размножения устроены сложно. Все трематоды гермафродиты, лишь немногие виды кровяных сосальщиков раздельнополы. Муж-
79
ские половые органы представлены двумя семенниками и отходящими от них семявыносящими протоками, образующими семяизвергательный канал, который заканчивается копулятивным органом — циррусом (см. рис. 47), находящимся в половой клоаке.
Женские половые органы представлены одним яичником, от которого начинается яйцевод, впадающий в оотип. Сюда же впадают протоки желез: желточных и тельца Мелиса. Оотип окружен мелкими скорлуповыми железами. От оотипа начинается длинная извитая матка, конец которой открывается женским половым отверстием рядом с мужским половым отверстием. Сосальщикам свойственно перекрестное оплодотворение, самооплодотворение возможно, но очень редко. Сперма с помощью копулятивного органа вводится в свою матку или матку другого сосальщика. По матке спермии поступают в семяприемник и накапливаются там. По мере необходимости спермии поступают в оотип. Сюда же выходят продукты придаточных желез и яйцеклетки из яйцевода. После оплодотворения яйца покрываются желточными клетками и затем прочной скорлуповой оболочкой. Оплодотворенные и покрытые скорлупой яйца поступают в матку и выводятся наружу. Яйцо имеет крышечку, открывающуюся при выходе из него личинки.
Развитие большинства сосальщиков протекает со сложными превращениями и со сменой хозяев. Первые промежуточные хозяева — всегда брюхоногие моллюски, пресноводные или наземные. Вторые промежуточные (дополнительные) хозяева (если они есть) — разные беспозвоночные и позвоночные животные. Есть виды сосальщиков, которые имеют трех промежуточных хозяев (рис. 48).
Наиболее опасны как паразиты сельскохозяйственных животных и человека следующие дигенетические сосальщики.
Печеночный сосальщик (Fasciola hepatica), или фасциола печеночная, имеет листовидное тело до 5 см длиной (см. рис. 47). На переднем конце тела расположено ротовое отверстие, окруженное ротовой присоской. Тегумент с шипиками. На брюшной стороне тела имеется брюшная присоска. Кишечник двуветвистый с множеством отростков. Два ветвистых семенника расположены в середине тела ниже компактного ветвистого яичника. По бокам тела находятся желточники.
Печеночный сосальщик паразитирует в желчных протоках печени растительноядных и всеядных животных, может поражать и человека, вызывая заболевание фасциолез. Питается сосальщик желчью. Сильнее всего поражаются овцы и молочный скот, которых пасут в поймах рек, на заливных лугах и т. п. Нередко болезнь может иметь летальный исход из-за закупоривания двуустками желчных протоков и невозможности оттока желчи из печени.
В желчных протоках паразиты копулируют, но возможно и самооплодотворение. С желчью оплодотворенные яйца, покрытые скорлуповыми оболочками, через кишечный тракт с калом хозяина попадают во внешнюю среду. За сутки один паразит может отложить сотни тысяч яиц (рис. 49).
80