

Гистология / Miadelets-OD_Gistologiia_tsitologiia_i_embriologiia_cheloveka_Ch-2_2016
.pdfских фолликулов с выраженной тека-тканью (см. ниже). Этот тип встречается наиболее часто.
в) Гиперпластический тип характеризуется активным ростом фолликулов (имеются многочисленные первичные фолликулы), кистозной атрезией многих из них на фоне гиперемии органа и гиперплазии интерстициальной железы.
Развитие яйцеводов, матки и влагалища. Мезонефральные протоки в женском организме подвергаются редукции. Однако до разрушения от них отделяются парные парамезонефральные (мюллеровы) протоки. Из верх-
них отделов этих протоков образуются парные яйцеводы, а нижние сливаются вместе, формируя матку и верхнюю часть влагалища. Нижняя часть влагалища образуется из мочеполового синуса.
Наружные половые органы развиваются из таких источников:
1)из полового бугорка формируется клитор;
2)из половых складок образуются малые половые губы;
3)из половых валиков развиваются большие половые губы. АНОМАЛИИ РАЗВИТИЯ МАТКИ И ВЛАГАЛИЩА (Рис. 24.2). При
нарушении эмбриогенеза матки и влагалища (нарушении слияния парамезонефральных протоков) может образоваться ряд патологических ситуаций:
седловидная, однорогая, двурогая матка, двойные влагалище и матка, и
т. д. Подобные аномалии могут приводить либо к невозможности развития беременности, либо к ненормальному положению плода (например, поперечное). В последнем случае самопроизвольные роды не наступают, и требуется кесарево сечение.
УСЛОВИЯ РАЗВИТИЯ ГОНАД И ДРУГИХ ПОЛОВЫХ ОРГАНОВ ПО ЖЕНСКОМУ ТИПУ
1. Отсутствие Y- хромосомы и андрогенов является определяющим фактором в развитии гонад по женскому типу. При этом индифферентная железа превращается в яичники. Напомним, что в мужском организме под действием мужских половых гормонов канальцы первичной почки превращаются в выносящие канальцы головки придатка. При отсутствии тестостерона и его производного дигидротестостерона вольфов проток и канальцы первичной почки подвергаются дегенерации (существует иная точка зрения, согласно которой канальцы мезонефроса участвуют в формировании сети яичника). Мочеполовой синус развивается в нижнюю часть влагалища, половой бугорок превращается в клитор, а половые складки и половые валики дифференцируются в малые и большие половые губы соответственно. Известно, что при приеме беременной женщиной андрогенов с лечебной целью в критический период (период дифференцировки половой системы, 8-я неделя эмбриогенеза) может возникнуть маскулинизация наружных половых органов.
451
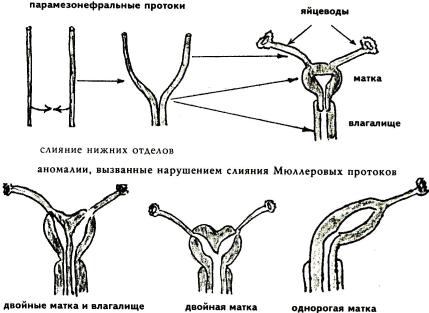
Рис. 24.2. Аномалии развития матки и влагалища
2. Действие эстрогенов. Они начинают синтезироваться фолликулярными клетками достаточно рано и способствуют
дифференцировке внегонадных органов женской половой системы: яйцеводов,
матки, влагалища.
3. Отсутствие мюллерова ингибирующего фактора (МИФ) ведет к тому, что мюллеров проток превращается в яйцеводы, матку и верхнюю часть влагалища.
ФУНКЦИОНАЛЬНАЯ МОРФОЛОГИЯ ЯИЧНИКОВ
ФУНКЦИИ ЯИЧНИКОВ. Яичники выполняют две основные функции: генеративную (образование женских половых клеток - яйцеклеток) и эндокринную - вырабатывают женские и мужские половые гормоны, а также ряд других гормонов и биологически активных веществ (о них см. ниже), регулирующих собственные функции яичников (внутрисистемный уровень регуляции).
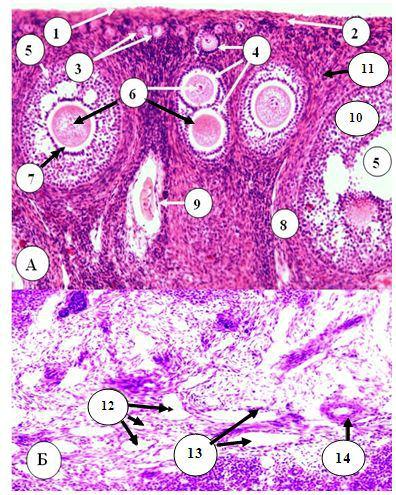
СТРОЕНИЕ. Яичники - парные паренхиматозные зональные органы. Их строму составляют белочная оболочка из плотной волокнистой соединительной ткани, веретеновидноклеточная соединительная ткань коркового вещества, в клеточном составе которой преобладают фибробласты и фиброциты, и рыхлая соединительная ткань мозгового вещества, В корковом веществе клетки располагаются характерно, формируя своеобразные завихрения. Снаружи от белочной оболочки находится видоизмененный мезотелий серозной оболочки, который обладает высокой пролиферативной активностью и очень часто является источником развития опухолей яичника. Паренхима яичников представлена совокупностью фолликулов и желтых тел, находящихся на разных стадиях развития (Рис. 24.3, 24.4).
Яичник подразделяется на корковое и мозговое вещество. В корковом веществе находятся примордиальные, первичные, вторичные, третичные и атретические фолликулы, а также желтые и беловатые тела. Мозговое вещество образовано РВНСТ, в которой находятся кровеносные сосуды, нервный аппарат, а также могут встречаться эпителиальные тяжи, представ-
452
ляющие собой остатки мезонефроса. Они могут быть источником развития кист яичника.
Рис. 24.3. Строение яичника А – корковое вещество: 1 – мезотелий яичника;
2 – белочная оболочка; 3 – примордиальные фолликулы; 4 – первичные фолликулы; 5 – третичный (везикулярный) фолликул яичника; 6 – первичный овоцит; 7 – прозрачная оболочка; 8 – тека фолликула; 9 – атретический фолликул; 10
– гранулеза (зернистый слой) фолликула; 11 – веретиновидноклеточная соединительная ткань стромы коркового вещества яичника;
Б – мозговое вещество яичника: 12 – вены; 13 – лимфатические сосуды; 14 - артерия
ЦИТОФИЗИОЛОГИЯ ОВАРИАЛЬНОГО ЦИКЛА
Овариальным циклом называют циклические изменения, происходящие в яичнике с наступлением полового созревания и направленные на образование женских половых клеток и гормонпродуцирующих структур. Он включает такие события:
1)рост фолликулов (фолликулогенез, фолликулиновая фаза);
2)овуляцию;
3)образование и функционирование желтого тела (лютеогенез, лю-
теиновая фаза). Овариальный цикл повторяется каждые 28-30 дней на протяжении репродуктивного периода женщины и в соответствии с происходящими процессами делится на 2 фазы: фолликулиновую и лютеиновую. Одним из важных моментов этого цикла является овуляция - разрыв зрелого фолликула и выход первичного овоцита в брюшную полость. Она приходится на середину овариального цикла и разграничивает две его фазы.
ФОЛЛИКУЛОГЕНЕЗ. Начинается с примордиальных фолликулов - мелких
453
фолликулов, расположенных непосредственно под белочной оболочкой. Они представляют собой первичный овоцит, окруженный одним слоем плоских фолликулярных клеток, лежащих на базальной мембране. Овоцит находится в диплотене профазы мейоза I и имеет достаточно крупное с дисперсным хроматином ядро, крупное ядрышко и слабо выраженные органеллы. Блокаду развития фолликула на этой стадии осуществляют фолликуляр-
ные клетки путем выработки ингибина. Таким образом, в приморди-
альном фолликуле первичный овоцит на длительные годы заторможен в состоянии мейоза и поэтому весьма чувствителен к различным повреждающим факторам, особенно к ионизирующей радиации. В связи с этим с возрастом у женщин в овоцитах накапливаются мутации, что потенциально может привести к врожденным уродствам и аномалиям развития потомства.
С наступлением большого роста овоцитов, который происходит в менструальном цикле у взрослой женщины, один из овоцитов, входящий в так называемый доминантный фолликул, увеличивается в размерах, а фолликулярные клетки становятся кубическими, затем призматическими. Доминантным называется фолликул, которому отдается предпочтение в развитии в данном менструальном цикле; фолликулярные клетки такого фолликула имеют наибольшее количество рецепторов к фоллитропину. В результате его развития образуются первичные фолликулы. Их отличительной особенностью являются следующие: 1) в них вокруг овоцита формируется прозрачная оболочка (zona pelucida, ZP); 2)происходит увеличение высоты фолликулярных клеток до кубических и затем – столбчатых. Однако эпителий остается однослойным. Первичные фолликулы появляются в яичнике еще до полового созревания.
ZP образована гликопротеинами. Она выполняет ряд функций: 1) участвует в формировании гемато-овариального барьера (см. ниже); 2) препятствует полиспермии; 3) обеспечивает видоспецифичность оплодотворения, а также 4) защищает эмбрион от неблагоприятных воздействий при прохождении его по половому тракту к месту имплантации в матке; 5) препятствует адгезии в яйцеводах и 6) обеспечивает компактное расположение бластомеров. Синтез ZP осуществляется овоцитом. Фолликулярные клетки не принимают в ее биосинтезе участия, но побуждают к нему овоцит, выделяя регуляторные вещества (существует также иная точка зрения, что участие в образовании ZP принимают и фолликулярные клетки). О начале синтеза ZP свидетельствует образование микроворсинок плазмолеммами контактирующих фолликулярных клеток и овоцита.
При дальнейшем развитии фолликула, происходящем только после полового созревания, фолликулярные клетки под действием фолликулостимулирующего гормона размножаются и гипертрофируются, в них появляется большое количество органелл и трофических включений. Фолликулярный эпителий становится многослойным, а фолликул – вторичным (Рис. 24.4,
454
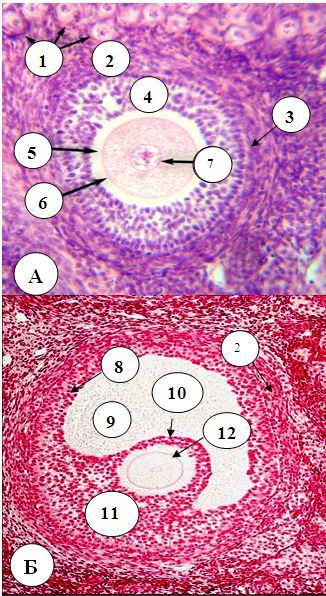
А). Фолликулярные клетки проникают отростками в отверстия в ZP и вступают в тесный контакт с овоцитом. Из окружающей соединительной ткани вокруг вторичных фолликулов формируется соединительнотканная оболочка
- тека фолликула.
Рис. 24.4. Строение яичника А – вторичный, Б – третичный фолликулы
1 – примордиальные фолликулы; 2 – тека фолликула; 3 – базальная мембрана фолликулярных клеток; 4 - многослойный кубический фолликулярный эпителий; 5 – прозрачная оболочка; 6 – плазмолемма первичного овоцита; 7 – ядро первичного овоцита; 8 – гранулеза (зернистый слой из фолликулярных клеток); 9 – полость третичного фолликула; 10 – лучистый венец; 11 – яйценосный бугорок; 12 – первичный овоцит
Тека состоит из двух слоев:
1. Наружного фиброзного слоя (фиброзная тека), пред-
ставленного плотной соединительной тканью и без резких границ переходящего в окружающую фолликул соединительную ткань. В нем содержатся миофибробласты, вызывающие сжатие фолликула после овуляции.
2. Внутреннего сосудистого слоя (сосудистая тека), образованного РСТ. Во внутренней теке разрас-
таются кровеносные сосуды и в большом количестве появляются эндокри-
ноциты (эндокринные интерстициальные клетки). Рост сосудов во внутренней теке стимулируется ангиогенным фактором, который синтезируется фолликулярными клетками. Таким образом, для вторичного фолликула характерны многослойная оболочка из фолликулярных клеток и тека
(Рис. 24.4, А).
В дальнейшем фолликулярные клетки образуют жидкость, которая формирует между фолликулярными клетками несколько полостей. Такие фолликулы называются третичными, антральными (рис. 24.4, Б). Овоцит в третичном фолликуле уже не увеличивается в размерах. Рост фолликула
455
происходит за счет увеличения размеров, количества фолликулярных клеток и последовательного накопления в полостях жидкости. Постепенно полости сливаются в одну, и овоцит вместе с покрывающими его фолликулярными клетками смещается на один полюс. В это время происходит завершение мейоза I, и первичный овоцит превращается во вторичный овоцит. Стенку фолликула образует многослойный фолликулярный эпителий, или тека гранулеза (зернистый слой), лежащий на базальной мембране. Смещенный к одному из полюсов первичный овоцит окружают фолликулярные клетки, формирующие яйценосный бугорок. Фолликулярные клетки, непосредственно окружающие овоцит, при помощи отростков тесно взаимодействуют с его плазмолеммой. Вместе со своими отростками эти клетки называются лучистым венцом. Таким образом, с образованием третичного фолликула фолликулярные клетки дифференцируются на два вида: клетки гранулезного слоя и клетки лучистого венца. Такой фолликул называется зрелым (предо-
вуляторным), или граафовым пузырьком.
Фолликулярные клетки выполняют ряд функций.
1.Трофическая функция заключается в передаче овоциту, удаленному от сосудов, питательных веществ.
2.Барьерно-защитная функция. Фолликулярные клетки участвуют в формировании гемато - овариального барьера (ГОБ), защищающего ово-
цит от аутоиммунных реакций организма и ряда токсических веществ, а также облегчающего поступление к овоцитам трофических и регуляторных веществ.
Всостав гемато-овариального барьера входят:
1.Эндотелий капилляров непрерывного типа;
2.Базальная мембрана эндотелиоцитов непрерывного типа с перицитами, способными к фагоцитозу;
3.Прослойка РВНСТ с фагоцитирующими макрофагами;
4.Базальная мембрана фолликулярных клеток;
5.Фолликулярные клетки;
6.Блестящая зона.
3.Секреторная функция. Фолликулярные клетки секретируют фолликулярную жидкость.
4.Фагоцитарная функция. При атрезии фолликула фолликулярные клетки фагоцитируют его остатки.
5.Эндокринные функции фолликулярных клеток достаточно обшир-
ны.
а) клетки участвуют в продукции эстрогенов, релаксина и прогестеро-
на. Последние два гормона образуются фолликулярными клетками после их трансформации в лютеоциты. Эстрогены образуются путем ароматизации из андрогенов, продуцируемых эндокриноцитами внутренней теки;
б) фолликулярные клетки крупных фолликулов продуцируют ингибин.
456
Ингибин блокирует выброс аденогипофизом фоллитропина, и, следовательно, мейоз других овоцитов, останавливая его в профазе первого деления. Так происходит ауторегуляция яичника на внутрисистемном уровне;
в) синтезируют простагландины, которые регулируют кровоснабжение фолликула;
г) секретируют фактор, аналогичный гонадолиберинам гипоталамуса д) секретируют ангиогенный фактор.
ОВУЛЯЦИЯ. СУЩНОСТЬ И МЕХАНИЗМЫ ОВУЛЯЦИИ. Овуляция представляет собой процесс разрыва третичного фолликула с выходом окруженного фолликулярными клетками овоцита II порядка в полость брюшины. Механизмы овуляции достаточно сложные и до конца не исследованы, но хорошо известно, что этот процесс опосредован рядом гормонов. Начало овуляции связано с резким выбросом в кровь гормона гипофиза лютропина. Этот гормон вызывает резкую гиперемию и отек яичника, что способствует диапедезу лейкоцитов, участвующих в разрушении стенки фолликула. Гиперемии способствуют и синтезируемые фолликулом кинины, цитокины. Возникает своеобразная воспалительная реакция в стенке фолликула. Кроме того, одновременно под действием лютропина в фолликулярных клетках вырабатываются простагландины Е и F, которые стимулируют активность протеаз и коллагеназы в гранулезных клетках и в нейтрофилах. Эти ферменты накапливаются в лизосомах клеток перед овуляцией. В это же время в фолликулярной жидкости накапливаются и ферменты, разрушающие гликозаминогликаны межклеточного вещества. Действие всех указанных ферментов ведет к истончению и ослаблению стенки фолликула. Продукты распада межклеточного вещества активируют воспалительную реакцию, действуя как хемоаттрактанты для все новых порций нейтрофилов, ферменты которых участвуют в разрушении стенки яичника над граафовым пузырьком. Параллельно происходит нарушение контактов между фолликулярными клетками гранулезной оболочки. Одновременно вокруг фолликула концентрируются миоидные клетки (по другим сведениям - гладкие миоциты), сокращение которых способствует разрыву фолликула. Сокращение этих клеток и миоцитов сосудов яичника вызывает окситоцин, который выбрасывается в большом количестве из задней доли гипофиза и поддерживает давление в полости фолликула на постоянном уровне, Играет роль также прекращение непосредственно перед овуляцией кровообращения в тканях яичника, расположенных над граафовым пузырьком, что ведет к их некрозу.
Происходят изменения и в микроциркуляторном русле третичного фолликула. Нарушаются межэндотелиальные контакты, эндотелиоциты начинают осуществлять интенсивный пиноцитоз. Нарастает отек стромы в окружении граафова пузырька.
В результате комплексного действия всех этих факторов белочная оболочка и покрывающий мезотелий истончаются и разрыхляются, формируя
457
так называемую стигму. Накануне овуляции разрушаются связи фолликулярных клеток лучистого венца с остальными клетками яйценосного бугорка, овоцит с клетками лучистого венца отделяется от бугорка и поступает в фолликулярную жидкость. После разрыва стигмы вторичный овоцит, окруженный фолликулярными клетками, выбрасывается в брюшную полость, захватывается фимбриями яйцеводов и движется в сторону полости матки. После контакта со сперматозоидами он претерпевает второе деление мейоза и превращается в зрелую овотиду и затем в зиготу.
Обычно овуляция протекает в одном третичном фолликуле, однако возможны случаи развития нескольких доминантных фолликулов, образование нескольких граафовых пузырьков и их овуляция. Это приводит к развитию разнояйцевых близнецов. Множественную овуляцию можно вызвать искусственно введением гонадотропинов, что применяют для экстракорпорального (in vitro) оплодотворения с дальнейшей имплантацией развивающегося эмбриона в полость матки. Экстракорпоральное оплодотворение используется при невозможности наступления беременности естественным путем. Последнее может иметь место, например, при воспалительных процессах в яйцеводах, приводящих к нарушению их проходимости. Напротив, введение эстрогенов и прогестерона подавляет рост фолликулов и овуляцию,
что используется в контрацепции (пероральные гормональные контрацептивы).
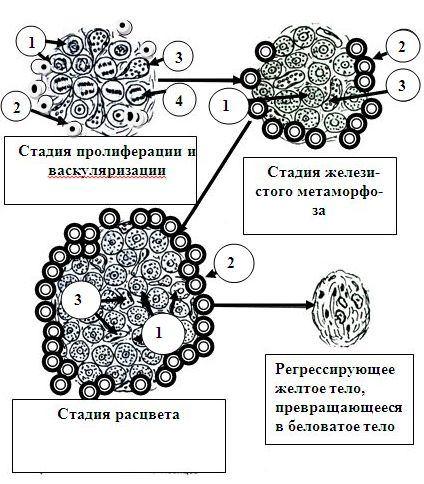
ЛЮТЕОГЕНЕЗ. СТАДИИ РАЗВИТИЯ ЖЕЛТОГО ТЕЛА. В лютеогенезе выделяют ряд стадий. Четыре из них отражены на рисунке 24.5.
1.Стадия геморрагического (красного) тела. На месте лопнувшего граафова пузырька остаются зернистый слой фолликулярных (гранулезных) клеток и тека фолликула, а в полость изливается кровь, которая быстро организуется (прорастает соединительной тканью).
2.Стадия пролиферации и васкуляризации. В эту стадию происходит размножение фолликулярных клеток и клеток теки, а также врастание между ними сосудов из сосудистой теки. Между текой и гранулезными клетками исчезает базальная мембрана.
3.Стадия железистого метаморфозa. Фолликулярные клетки зерни-
стого слоя превращаются в крупные полигональные железистые (лютеиновые) клетки: в них резко увеличивается объем гладкой эндоплазматической сети, гипертрофируется комплекс Гольджи, накапливается пигмент лютеин
ижировые (холестериновые) включения, появляются митохондрии с крупными кристами. Это так называемые гранулезные лютеоциты. Интерстициальные клетки внутреннего слоя теки превращаются в лютеоциты теки (тека-лютеоциты). Они имеют меньшие размеры и окрашиваются более интенсивно, чем гранулярные лютеоциты. Тека-лютеоциты находятся на периферии желтого тела и продуцируют мужские половые гормоны. В результате интенсивного разрастания лютеоцитов и прорастания соединительной ткани
458
полость бывшего третичного фолликула исчезает.
4. Стадия расцвета. Лютеоциты начинают продуцировать гормон прогестерон, который расслабляет мускулатуру матки, приводит к увеличению толщины ее слизистой оболочки, активирует секрецию маточных желез. В желтом теле вырабатывается также гормон белковой природы релаксин, который, как и прогестерон, расслабляет мускулатуру матки, а также (за счет накопления цАМФ) активирует ферменты в хондроцитах лонного сочленения. Они разрушают часть компонентов лонной связки, которая при этом легко поддается растяжению, что ведет к расхождению костей и увеличению полости таза. Таким образом, два гормона желтого тела готовят организм к беременности и обеспечивают ее нормальное протекание. Синтез релаксина резко усиливается в конце беременности. Кроме того, прогестерон тормозит образование новых фолликулов в яичнике и предотвращает наступление повторной беременности. Желтое тело вырабатывает также эстрогены, андрогены (некоторое количество их продуцируется тека-лютеиновыми клетками), окситоцин и простагландины.
Рис. 24.5. Развитие желтого тела (стадия геморрагического тела не отражена)
- стадия пролиферации и васкуляриза-
ции: 1, 4 – делящиеся фолликулярные клетки; 2 – интерстициальные клетки внутренней теки; 3 – кровеносный капилляр; - стадия железистого
метаморфоза: 1 – гра-
нулезные лютеоциты; 2
– тека-лютеоциты; 3 – кровеносный капилляр; - стадия расцвета: 1 –
гранулезные лютеоциты; 2 – текалютеоциты; 3 – кровеносный капилляр
5. Стадия регресса желтого тела. Лютеиновые клетки уменьшаются в размерах и атрофируются. Накопление в клетках холестерина, не используемого для синтеза прогестерона, ведет к гибели лютеоцитов и замещению их
459
соединительной тканью. В лютеолизе большую роль играют также простагландины, которые в фазу регресса локально вырабатываются в самом желтом теле, а также в матке. Эндокринная функция желтого тела прекращается. В конечном итоге желтое тело прорастает соединительной тканью и превращается в беловатое тело. Продолжительность существования желтого тела зависит от того, наступило или нет оплодотворение. Если оно не произошло и беременность не наступила, то стадия расцвета желтого тела длится около 10-12 дней. Такое желтое тело называется менструальным желтым телом. При наступлении беременности желтое тело функционирует на протяжении почти всей беременности, подавляя рост новых фолликулов, предотвращая повторные оплодотворения и выкидыш. Это желтое тело называется желтым телом беременности. Разница между двумя видами желтых тел заключается лишь в размерах и времени функционирования: желтое тело беременности крупнее, достигает размеров до 3 см и функционирует дольше, чем менструальное желтое тело, существующее в течение 6 мес беременности. Поскольку обратное развитие желтых тел идет медленнее, чем их образование, то в яичнике можно увидеть до пяти желтых тел на разных стадиях регрессии. Регрессией желтого тела заканчивается овариальный цикл.
АТРЕЗИЯ ФОЛЛИКУЛОВ. Стадии третичного фолликула достигают не все фолликулы, и в течение репродуктивной жизни женщины в яичниках образуется лишь около 500 яйцеклеток. Все остальные фолликулы на разных стадиях своего развития подвергаются разрушению - атрезии. Особенно много фолликулов разрушается в период от рождения до 18 лет, всего же атрезии подвергаются более 99% фолликулов, имеющихся к моменту рождения.
Различают дегенеративный и текогенный типы атрезии. При дегене-
ративном типе атрезии вначале происходит разрушение овоцита. Фолликулярные клетки некоторое время сохраняются, формируя микрокисту, которая впоследствии прорастает соединительной тканью, а затем бесследно исчезает. Дегенеративной атрезии подвергаются примордиальные и первичные фолликулы. Она всегда множественная, т.е. разрушаются целые группы фолликулов.
Текогенной атрезии подвергаются вторичные и третичные фолликулы. Эта атрезия, наоборот, начинается с гибели фолликулярных клеток, и лишь затем гибнет овоцит. Изменения начинаются с исчезновения микроворсинок на поверхности фолликулярных клеток и потери их связи с овоцитом. Далее фолликулярные клетки отслаиваются от базальной мембраны. Нарастает интенсивность апоптоза этих клеток. Лишь после того, как процессы гибели фолликулярных клеток достигают значительного объема, начинаются дегенеративные изменения овоцита. Дольше всех сохраняется оксифильная блестящая оболочка, по которой на микропрепаратах и узнают атретические
460