

31. Строение и значение мерцательных ресничек.
Однослойный многорядный призматический реснитчатый (мерцательный) эпителий воздухоносных путейнаиболее типичный представитель многорядных эпителиев. В нем имеются клетки четырех основных типов: (1) низкие вставочные (базальные), (2) высокие вставочные (промежуточные), (3) реснитчатые (мерцательные) и (4) бокаловидные. Камбиальными элементами служат низкие вставочные клетки, которые имеют мелкие размеры. Своим широким основанием они прилежат к базальной мембране, обусловливая прикрепление к ней всего эпителиального пласта, а узкой апикальной частью не доходят до просвета. Наиболее дифференцированные клетки эпителия — реснитчатые (численно преобладают) и бокаловидные. Последние вырабатывают слизь, которая покрывает поверхность эпителия, перемещаясь по ней благодаря биению ресничек мерцательных клеток. Реснитчатые и бокаловидные клетки своей узкой базальной частью контактируют с базальной мембраной и прикрепляются к вставочным клеткам, а апикальной — граничат с просветом органа.
32. Строение и значение микроворсинок.
Микроворсинки — пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. Микроворсинки обеспечивают многократное увеличение площади поверхности клетки, на которой происходит расщепление и всасывание веществ. На апикальной поверхности некоторых клеток, активно участвующих в указанных процессах (в эпителии тонкой кишки и почечных канальцев) имеется до нескольких тысяч микроворсинок, образующих в совокупности щеточную каемку.
Каркас
каждой микроворсинки образован пучком,
содержащим около 40 микрофиламентов,
лежащих вдоль ее длинной оси. В апикальной
части микроворсинки этот пучок закреплен
в аморфном веществе. Его жесткость
обусловлена поперечными сшивками из
белков фимбрина и виллина, изнутри пучок
прикреплен к плазмолемме микроворсинки
особыми белковыми мостиками (молекулами
минимиозина). У основания микроворсинки
микрофиламенты пучка вплетается в
терминальную сеть, среди элементов
которой имеются миозиновые филаменты.
Взаимодействие актиновых и миозиновых
филаментов терминальной сети, вероятно,
обусловливает тонус и конфигурацию
микроворсинки
33-37
38.

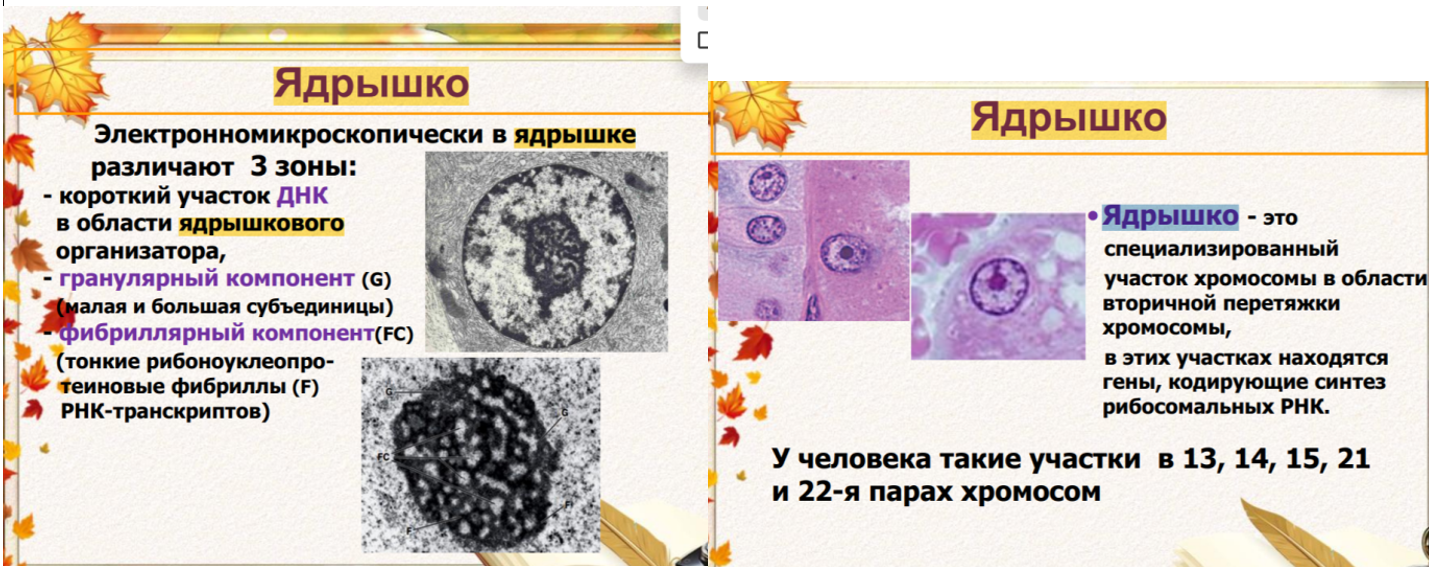 39.
39.
 40.
40.

 41.
41.
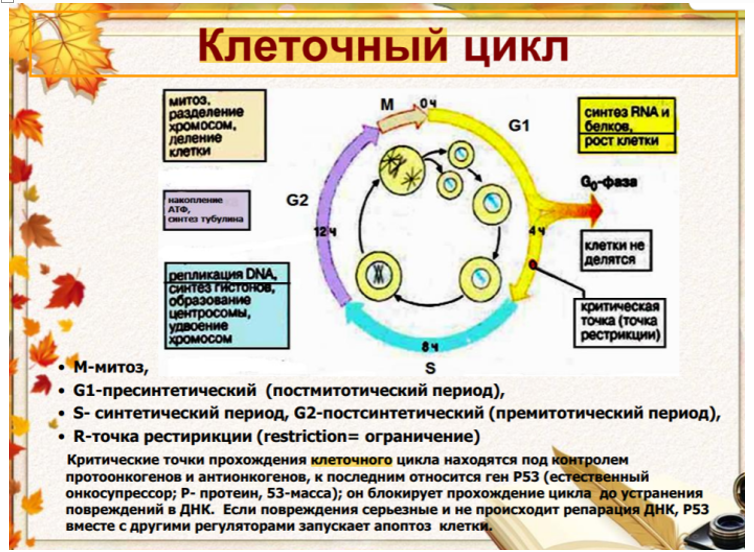
 42.
Синтетический (S) период. Его значение.
42.
Синтетический (S) период. Его значение.
1. Синтетический (S-) период характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов.(2n4c)
43. Премитотический (G2) период, происходящие в нём события.
Постсинтетический или премитотический (G2-) период следует за S-периодом и продолжается вплоть до митоза (часто обозначаемого буквой М). В течение этого периода клетка осуществляет непосредственную подготовку к делению. Происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (в частности, тубулин), необходимые для процесса деления. Длительность G2-периода составляет 2-4 часа. Возможность выхода клетки из G2-периода в G0-период с последующим возвращением в G2-период в настоящее время большинством авторов отрицается.
Контроль вступления клетки в митоз осуществляется двумя специальными факторами с противоположно направленными эффектами: митоз тормозится до момента завершения репликации ДНК М-задерживающим фактором и индуцируется М-стимулирующим фактором. Действие последнего проявляется лишь в присутствии других белков — циклинов (синтезируются на протяжении всего цикла и распадаются в середине митоза).(2n4c)
44. Митотический (М) период, его общая характеристика, стадии. Митотический период или Митоз (от греч. mitos — нить), называемый также кариокинезом, или непрямым делением клеток, является универ- сальным механизмом деления клеток. Митоз следует за G2—периодом и завершает клеточный цикл. Он длится 1-3 часа и обеспечивает равномерное распределение генетического материала в дочерние клетки. Митоз включает 4 основные фазы: профазу, метафазу, анафазу и телофазу.
45. Характеристика стадий митоза: профазы, метафазы, анафазы, телофазы.
Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядрышко и ядерная оболочка к концу фазы исчезают (последняя распадается на мембранные пузырьки, сходные с элементами ЭПС, а поровый комплекс и ламина диссоциируют на субъединицы. Кариоплазма смешивается с цитоплазмой. Центриоли мигрируют к противоположным полюсам клетки и дают начало нитям митотического (ахроматинового) веретена. В области центромеры образуются особые белковые комплексы — кинетохоры, к которым прикрепляются некоторые микротрубочки веретена (кинетохорные микротрубочки); показано, что кинетохоры сами способны индуцировать сборку микротрубочек и поэтому могут служить центрами организации микротрубочек. Остальные микротрубочки веретена называются полюсными, так как они протягиваются от одного полюса клетки к другому; лежащие вне веретена микро- трубочки, расходящиеся радиально от клеточных центров к плазмолемме, получили наименование астральных или микро- трубочек (нитей) сияния.(2n4c)
Метафаза соответствует максимальному уровню конденсации хромосом, которые выстраиваются в области экватора митотического веретена, образуя картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со стороны полюсов). Хромосомы перемещаются в экваториальную плоскость и удерживаются в ней благодаря сбалансированному натяжению кинетохорных микротрубочек. Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.(2n4c)
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды
(в области центромеры) и движения дочерних хромосом к противоположным полюсам клетки, которое происходит вдоль микротрубочек веретена со скоростью 0.2-0.5 мкм/мин. Сигнал к началу анафазы включает резкое (на порядок) повышение концентрации Са2+ в гиало- плазме, выделяемого мембранными пузырьками, образующими скопления у полюсов веретена. Механизм движения хромосом в анафазе окончательно не выяснен, однако установлено, что в области веретена помимо актина имеются такие белки как миозин и динеин, а также ряд регуляторных белков и Са2+-АТФаза. По некоторым наблюдениям, оно обусловлено укорочением (разборкой) микротрубочек, прикрепленных к кинетохорам. Анафаза характеризуется удлинением митотического веретена за счет некоторого расхождения полюсов клетки. Она завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картины звезд (стадия дочерних звезд). В конце анафазы благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки (сократимое кольцо), начинает образовываться клеточная перетяжка, которая углубляясь, в следующей фазе приведет к цитотомии.(4n4c всего или 2n2c у каждого полюса)
Телофаза — конечная стадия митоза, в течение которой реконструируются ядра дочерних клеток и завершается их разделение. Вокруг конденсированных хромосом дочерних клеток из мембранных пузырьков (по другим данным, из аЭПС) восстанавливается кариолемма, с которой связывается формирующаяся ламина, вновь появляются ядрышки, которые образуются из участков соответствующих хромосом. Ядра клеток постепенно увеличиваются, а хромосомы прогрессивно деспирализуются и исчезают, замещаясь картиной хроматина интерфазного ядpa. Одновременно происходит углубление клеточной перетяжки, и клетки в течение некоторого времени остаются связанными суживающимся цитоплазматическим мости- ком, содержащим пучок микротрубочек (срединное тельце). Дальнейшая перешнуровка цитоплазмы завершается формированием двух дочерних клеток. В телофазе происходит распределение органелл между дочерними клетками; равномерности этого процесса способствует то, что одни органеллы достаточно многочисленны (например, митохондрии), другие (подобно ЭПС и комплексу Гольджи) во время митоза распадаются на мелкие фрагменты и пузырьки.(2n2c) 46-48 49. 49 Протоонкогены – группа генов-активаторов, контролирующих нормальное клеточное деление и дифференцировку. Продукты экспрессии этих генов воздейтвуют на разные механизмы регуляции деления клетки: на уровне активирующего сигнала, его рецептора в мембране, второго посредника или транскрипции. К настоящему времени идентифицировано более 50 протоонкогенов.
Установлено, что изменения структуры и усиление активности экспрессии протоонкогенов вызывает развитие опухолей. Повышние активности протоонкогенов может быть связано с изменением строения ДНК (в результате мутаций), увеличением количества генов (генной амплификации) или их реаранжировки, про которо гены размещаются вблизи активного промотора. Изменённые мутацией, но активные формы протоонкогенов носят название онкогенов. Злокачественная трансформация клтки может возникнуть не только вследствии повышения активности протоонкогенов, но и в результат снижения активности другой группы генов, называемых антионкогенами.
Антионкогены – гены, продукты которых – супрессоры опухолевого роста – угнетают митотическую активность клток. Из них наиболее подробно изучены гены RB, DCC, APC, WTI, NFI и особенно ген р53.
Инактивация функции антионкогенов и развитие опухолей . Потеря функции гена р53 (в результате мута- ции или делении) приводит к утрате контроля над клеточным циклом: клеткимутанты продолжают активно пролифериро- вать, несмотря на повреждения ДНК. Выявлена четкая связь между утратой функции гена р53 и развитием более 50 видов злокачественных опухолей у человека. Так, изменения гена р53 обнаружены в 55-70% случаев рака легкого, в 25-30% ра- ка молочной железы. Опухоли с потерей функции гена р53 характеризуются наиболее злокачественным течением. В некото- рых видах опухолей (в 60% меланом и лейкозов, в 80% глиом) обнаруживаются изменения гена р16; описаны опухоли, свя- занные с дефектами гена р15. Клетки рака шейки матки часто содержат инактивированные гены RB и р53. Мутация гена RB обнаруживается при ретинобластоме, опухолях костей, мочевого пузыря, легкого и молочной железы. Деления гена DCC характерна для опухолей толстой и прямой кишки, АРС для аденоматозного полипоза толстой кишки.
Факторы роста являются важными стимуляторами клеточного деления. Они представляют собой белки, усиливаю- щие митотическую активность в определенных тканях (тканяхмишенях). Их действие опосредуется специфическими ре- цепторами на плазмолемме клеток. К ним относятсяфактор роста нервов (ФРН),эпидермальный фактор роста, (ЭФР), тромбоцитарный фактор роста (ТРФР), инсулиноподобные факторы роста (ИФР), фактор роста фибробластов (ФРФ), колоше стимулирующие факторы (КСФ) стимуляторы отдельных этапов гематопоэза, интерлейкины (ИЛ) -1, -2 и -3. Список факторов роста постоянно расширяется. Высказывается предположение, что большинство типов клеток реагирует не на один специфический фактор роста, а на их комбинации. Некоторые факторы роста циркулируют в крови, но большинство действует в тканях локально (паракринно). Описаны также факторы, подавляющие клеточное деление.
Кейлоны (от греч. chalao успокаивать) представляют собой класс гормоноподобных регуляторов, угнетающих кле- точное размножение. Они являются полипептидами или гликопротеинами и обладают тканевой и клеточной специфично- стью. Кейлоны образуются всеми зрелыми дифференцированными клетками и
локально воздействуют на незрелые клетки этой же ткани, способные к делению. Они обеспечивают гомеостаз численности клеточной популяции, а их выделение кон- тролируется механизмом отрицательной обратной связи. Уменьшение численности популяции клеток (например, потеря клеток эпидермиса при ранении или лейкоцитов при кровотечении) вызывает снижение ингибирующего воздействия кейло- нов и подъем митотической активности в соответствующей ткани. Кейлоны участвуют в регуляции роста тканей, заживле- ния ран, иммунных реакций и других процессах.
50 Некроз (от греч. nekrosis умирание) возникает под действием резко выраженных повреждающих факторов пе- регревания (гипертермии), переохлаждения (гипотермии), недостатка кислорода (гипоксии), нарушения кровоснабжения (ишемии), метаболических ядов, химических препаратов, механической травмы и др. Некроз представляет собой "смерть в результате несчастного случая" и часто охватывает различные по численности группы клеток.
Структурно-функциональные изменения клеток при некрозе на начальных этапах его развития проявляются набуха- нием цитоплазмы и отдельных органелл (в особенности, митохондрий). Отмечается дисперсия рибосом, расширение цистерн ЭПС. Эти морфологические изменения обусловлены нарушением избирательной проницаемости плазмолеммы и развивают- ся в ответ на прекращение деятельности мембранных ионных насосов (изза непосредственного повреждения мембраны или вследствие отсутствия необходимой энергии). Повышение концентрации Са 2+ в гиалоплазме вызывает активацию свя- занных с мембраной фосфолипаз, которые разрушают мембранные фосфолипиды и вызывают обширные повреждения мем- бран. Разрушение клеточных структур резко ускоряется на поздних стадиях некроза после выделения гидролаз и других ферментов из поврежденных лизосом.
Изменения ядра при некрозе связаны с расщеплением ядерной ДНК лизосомальной ДНКазой на фрагменты различной длины (без какойлибо закономерности). Первоначально гетерохроматин конденсируется в виде крупных глыбок под ка- риолеммой, однако он не образует четко очерченных скоплений полулунной формы, которые характерны для ядер клеток, подвергающихся апоптозу (см. ниже). В дальнейшем ядро уменьшается, уплотняется (явление кариопикноза от греч. karyon ядро и pycnosis уплотнение греч.), распадается (подвергается кариорексису от греч. karyon ядро и rhexis
— разрыв) и лизируется (явление кариолизиса от греч. karyon ядро и lysis разрыв).
Поздние явления при некрозе включают разрыв ядерной оболочки, плазмолеммы и мембран органелл, разрушение и растворение ядра, утрату базофилии набухшей цитоплазмой, исчезновение клеточных границ и распад клетки.
Для некроза, в отличие от апоптоза не являющегося активным процессом (см. ниже), не требуется продолжающейся синтетической активности клетки, он не сопровождается активацией путей внутриклеточной сигнализации. Продукты рас- пада клеток попадают в межклеточные пространства, привлекают лейкоциты и макрофаги, фагоцитирующие клеточный детрит. Фагоциты, в свою очередь, выделяют разнообразные вещества, которые обусловливают активацию и приток раз- личных клеток вследствие хемотаксиса. Описанным образом развивается и в течение определенного времени поддержива- ется воспалительная реакция на продукты разрушения клеток при их некрозе.