

Средства, влияющие на афферентную иннервацию
Местноанестезирующие вещества. Классификация. Механизм анестезирующего действия и его локализация. Последовательность выключения разных видов чувствительности.
Виды местной анестезии. Сравнительная оценка современных анестетиков и их применение при разных видах анестезии. Токсическое действие анестезирующих веществ. Меры по его предупреждению.
Лекарственные средства, понижающие чувствительность окончаний афферентных нервов или препятствующие их возбуждению (вяжущие, обволакивающие и адсорбирующие средства). Показания к применению.
Лекарственные средства, стимулирующие окончания афферентных нервов (раздражающие средства, стимуляторы дыхания рефлекторного действия, горечи, слабительные и желчегонные средства рефлекторного действия). Показания к применению.
Нейротропные средства
Нейротропные средства. Определение. Общие принципы строения эфферентной нервной системы. Концепция химической передачи нервного возбуждения и организация химического синапса. “Мишени” для нейротропных средств.
Нейротропные средства - лекарственные соединения, реализующие свою основную фармакологическую активность за счет влияния на передачу нервного возбуждения в химическом синапсе. Усиливая, уменьшая или даже полностью блокируя передачу нервного импульса в синапсе, они могут менять функциональное состояние соответствующих органов и систем.
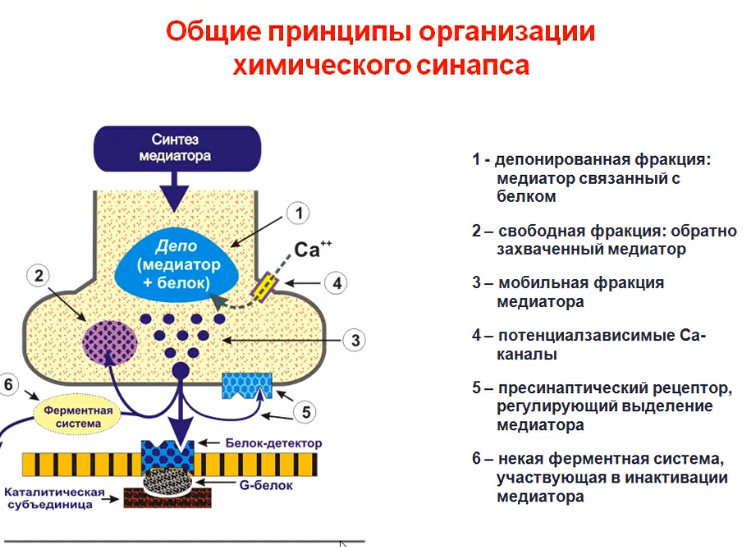
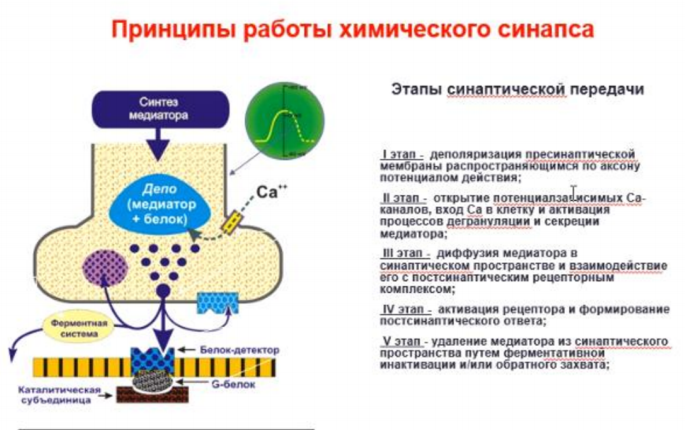
Обязательные компоненты химического синапса Наличие медиатора или эндогенного лиганда, т.е. химического передатчика нервного возбуждения. Обязательное существование высокоспецифичного для данного медиатора рецептора; как результат взаимодействия медиатор с рецептором, должен наблюдаться тот или иной физиологический эффект. Необходимым условием для любого синапса является наличие системы инактивации или прекращения взаимодействия в системе рецептор- медиатор. Последнее условие обязательно, поскольку синапс по существу динамическая система и превышение установленного временного лимита для существования комплекса лиганд- рецептор, переводит синапс в статический режим и, как результат, нарушение нормальной передачи возбуждения
“Мишени” для нейротропных средств. На пресинаптическом уровне различные БАВ могут влиять на процессы синтеза, депонирования и выделения медиатора, усиливая или тормозя эти процессы. Вещества могут прямо взаимодействовать с рецепторными комплексами, в результате этого они или имитируют эффекты медиатора или препятствуют взаимодействию медиатора с рецептором, т.е. могут вести себя как агонисты или антагонисты. Точкой приложения для некоторых фармакологических веществ на уровне синапса, могут быть системы инактивации медиатора. При этом вещества ослабляющие эти системы усиливают передачу нервного возбуждения в синапсе, а вещества повышающие активность процессов инактивации - ослабляют передачу в синапсе.
Структура и функционирование холинергического синапса. Структура холинергического рецептора. Классификация и локализация холинергических рецепторов.
Ацетилхолин - медиатор нервных импульсов в различных синапсах НС. Синтез происходит в аксоплазме холинергических нервов из активной уксусной кислоты и аминоспирта холина и катализируется ферментом холинацетилазой. Депонируется в связанной с белком форме и накапливается в непосредственной близости от пресинаптической мембраны Выброс происходит под действием приходящего нервного импульса, и спонтанно в виде квантов, содержащих несколько тысяч молекул медиатора. Спонтанное выделение медиатор занимает важное место в поддержании функциональной готовности мышцы. Существенную роль в процессе выделения медиатора играет деполяризация пресинаптической мембраны и вхождение в клетку ионов Са++ АХ играет роль гуморального фактора, участвуя в реализации внесинаптических эффектов. Он вызывает освобождение K+, связанных с белками, изменяет проницаемость биомембран, участвует в регуляции селективной проницаемости эритроцитов, регулирует активность некоторых дыхательных ферментов, влияет на метаболизм макроэргических фосфатных соединений и т.д. В основе немедиаторных эффектов АХ лежит его способностью изменять проницаемость мембран для ионов, особенно Са++
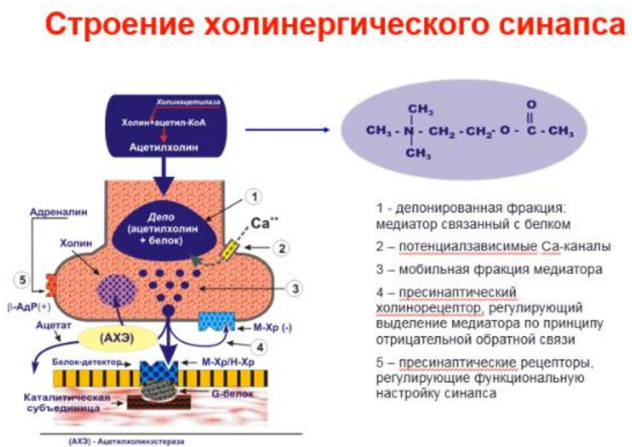
Холинорецептор - белковая макромолекула, способная избирательно взаимодействовать АХ, причем это взаимодействие вызывает краткотекущее изменение физико-химических свойств мембраны, в которую ХР встроен. Имеет два активных центра: 1. отрицательно заряженный анионный центр, в который входят карбоксильные группы глутамата или аспаргината; этот центр вступает во взаимодействие с положительной катионной головкой АХ; 2. эстерофильный центр, взаимодействующий с кислородом карбонильной группой или кислородом эфирной связи.
Молекула АХ содержит 3 пространственно разобщенных центра, имеющих значение для его биологической активности. АХ взаимодействует с холинорецептором с помощью двух центров, одним из которых обязательно является катионная головка. Вторым центром взаимодействия может быть «карбонильный» или «эфирный» кислород. В этом состоит отличие между М- и Н-холинорецепторами.
Предполагают тесную связь активации М-холинорецепторов с системой фосфотидилинозитола и гуанилатциклазы и повышение концентрации клеточного цГМФ, который является вторичным “мессенджером” в клетке. Это может быть следствием и того, что открываются связанные с рецептором Са++-инофоры (потенциалнезависимые каналы) и увеличивается внутриклеточная концентрация Са++
В настоящее время идентифицированы пресинаптические рецепторы, располагающиеся на пресинаптических окончаниях нервов. Пресинаптические М-холинорецепторы регулируют освобождение АХ по принципу отрицательной обратной связи, т.е. их стимуляция снижает высвобождение медиатора. Роль пресинаптических Н-рецепторов, обнаруженных в нервно-мышечном аппарате и ЦНС обратная, их стимуляция усиливает секрецию АХ.
Механизм гидролиза АХ. Молекула АХ является полярной: - триметиламмониевая группировка (катионная головка) - положительный заряд, - карбоксильная -отрицательный. В активном центре ХЭ также два участка, несущих заряд: - анионный - образованный аспаргиновой кислотой, - эстеразный пункт, в который входят серин, гистидин и тирозин. Катионная головка молекулы АХ фиксируется на анионном пункте активного центра холинэстеразы, а эстеразный пункт ХЭ образует ковалентную связь с углеродом карбоксильной группы АХ. Происходит разрыв эфирной связи в молекуле АХ с образованием свободного холина и ацетилированного фермента, который очень быстро регенерирует
М- и Н-холинорецепторы и их локализация
М-холинорецепторы |
Н-холинорецепторы |
расположены в постсинаптической мембране клеток эффекторных органов у окончаний постганглионарных холинергических (парасимпатических) волокон (гладкие мышцы, железы), в ЦНС (кора головного мозга, ретикулярная формация, лимбическая система, бульбарные центры, гипоталамус). Локализованы на постганглионарных окончаниях симпатических нервных волокон (потовые железы, гладкие мышцы сосудов нижних конечностей и органов малого таза). Установлена гетерогенность - М1-ХР - в вегетативных ганглиях и в ЦНС - М2-ХР - в сердце и на пресинаптической мембране холинергических синапсов и в сердце - М3-ХР - в гладких мышцах внутренних органов и экзокринных железах |
клетки ганглиев вег НС, хромафинные клетки надпочечников, хеморецепторы сосудов и ЦНС (кора, пирамидная система, спинной мозг), и нервно-мышечные синапсы. Существует гетерогенность разной локализации. Так фармакологическая чувствительность Н-ХР нервно-мышечных синапсов отличается Н-ХР вегетативной системы
|