

Коровкин биохимия
.pdfвый комплекс–гликокаликс. В поверхностных слоях гликокаликса задерживаются крупные молекулы и бактерии. Полисахариды не проникают через гликокаликс и, оставшись нерасщепленными при полостном пищеварении, гидролизуются на поверхности энтероцитов. Мальтоза, сахароза и лактоза могут гидролизоваться в гликокаликсе. Такое переваривание получило название пристеночного, или внеклеточного, пищеварения.
Маловероятным представляется всасывание значительных количеств дисахаридов, так как из экспериментов с парентеральным их введением известно, что большая часть дисахаридов, поступивших в кровяное русло, выделяется с мочой неизмененной; это является тем единственным и притом нефизиологическим случаем, когда дисахариды появляются в моче.
Скорость всасывания отдельных моносахаридов различна. Глюкоза и галактоза всасываются быстрее, чем другие моносахариды. Принято считать, что всасывание маннозы, ксилозы и арабинозы осуществляется преимущественно путем диффузии, всасывание же большинства других моносахаридов происходит за счет активного транспорта.
Щеточная каемка энтероцитов содержит системы переносчиков. Установлено существование переносчика, способного связывать различными своими участками глюкозу и Na+ и переносить их через плазматическую мембрану кишечной клетки. Считают, что глюкоза и Na+ высвобождаются затем в цитозоль, позволяя переносчику захватить новую порцию «груза». Na+ транспортируется по градиенту концентрации, стимулируя переносчик к транспорту глюкозы против указанного градиента. Свободная энергия, необходимая для этого активного транспорта, образуется благодаря гидролизу АТФ связанному с натриевым насосом, который «откачивает» из клетки Na+ в обмен на К+ . Динамика происходящих при этом процессов пока остается недостаточно ясной и в настоящее время обстоятельно изучается.
Судьба всосавшихся моносахаридов. Более 90% всосавшихся моносахаридов (главным образом глюкоза) через капилляры кишечных ворсинок попадает в кровеносную систему и с током крови через воротную вену доставляется прежде всего в печень. Остальное количество моносахаридов поступает по лимфатическим путям в венозную систему. В печени значительная часть всосавшейся глюкозы превращается в гликоген, который откладывается в печеночных клетках в форме своеобразных, видимых под микроскопом блестящих гранул.
СИНТЕЗ И РАСПАД ГЛИКОГЕНА
Гликоген–главная форма запасания углеводов у животных и человека. Накапливается гликоген главным образом в печени (до 6% от массы печени) и в скелетных мышцах, где его содержание редко превышает 1%. Запасы гликогена в скелетных мышцах ввиду значительно большей массы последних превышают его запасы в печени. Гликоген присутствует в цитозоле в форме гранул диаметром от 10 до 40 нм. На электронных микрофотографиях гликогеновые гранулы выглядят плотными. Установлено, что эти гранулы, кроме гликогена, содержат ферменты, катализирующие синтез и распад гликогена. Однако гликогеновые гранулы отличаются от мультиферментных комплексов (например, от пируватдегидрогеназного комплекса). Степень структурной организации гликогеновых гранул ниже, чем в мультиферментных комплексах. Следует подчеркнуть, что синтез и распад гликогена в клетке осуществляются разными метаболическими путями.
321
В частности, считалось, что гликогенфосфорилаза (фосфорилаза а) катализирует как распад гликогена, так и его синтез, потому что в опытах in vitro было показано, что гликогенфосфорилазная реакция обратима. Однако в дальнейшем было установлено, что в клетке (in vivo) фосфорилаза а катализирует только распад гликогена, синтез гликогена осуществляется при участии совершенно другого фермента. Оба эти процесса (синтез и распад гликогена) регулируют содержание глюкозы в крови и создают резерв глюкозы для интенсивной мышечной работы.
Синтез гликогена (гликогенез)
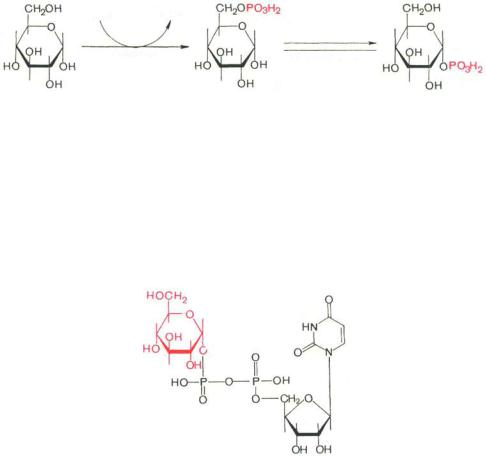
Прежде всего глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в печени–и глюкокиназы. Далее глюкозо-6-фосфат под влиянием фермента фосфоглюкомутазы переходит в глюкозо-1-фос- фат *:
АТФ |
АДФ |
|
|
Гексокиназа |
Фосфоглюкомутаза |
|
|
|
Глюкоза |
Глюнозо-6-фосфат |
Глюкозо-1-фосфат |
Образовавшийся глюкозо-1-фосфат уже непосредственно вовлекается в синтез гликогена. На первой стадии синтеза глюкозо-1-фосфат вступает во взаимодействие с УТФ (уридинтрифосфат), образуя уридиндифосфатглюкозу (УДФ-глюкоза) и пирофосфат. Данная реакция катализируется ферментом глюкозо-1-фосфат-уридилилтрансферазой (УДФГ-пирофосфорила- за):
Глюкозо-1-фосфат + УТФ <=> УДФ-глюкоза + Пирофосфат.
Приводим структурную формулу УДФ-глюкозы:
Уридиндифосфатглюноза (УДФ-глюкоза)
*Роль кофакторов в данной реакции выполняет глюкозо-1,6-бисфосфат, образующийся
вреакции, катализируемой фосфоглюкокиназой: глюкозо-1-фосфат + АТФ<=>глюкозо-1,6-бис- фосфат + АДФ.
322
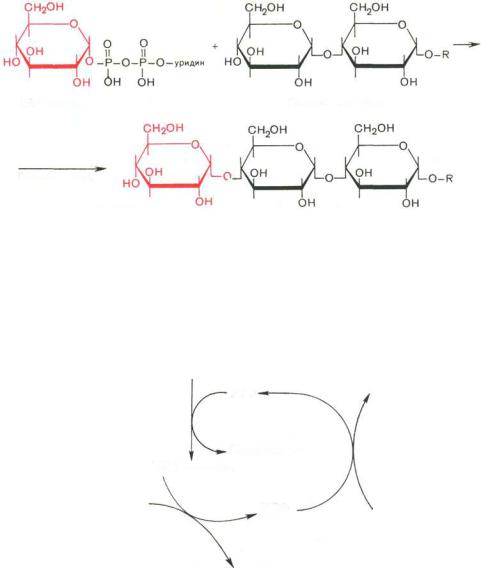
На второй стадии – стадии образования гликогена – происходит перенос глюкозного остатка, входящего в состав УДФ-глюкозы, на глюкозидную цепь гликогена («затравочное» количество). При этом образуется α-(1–>4)- связь между первым атомом углерода добавляемого остатка глюкозы и 4-гидроксильной группой остатка глюкозы цепи. Эта реакция катализируется ферментом гликогенсинтазой. Необходимо еще раз подчеркнуть, что реакция, катализируемая гликогенсинтазой, возможна только при условии, что полисахаридная цепь уже содержит более 4 остатков D-глю- козы.
УДФ-глюкоза |
Гликоген (n остатков) |
+ УДФ
Гликогенсинтаза
Гликоген (n+1 остатков)
Образующийся УДФ затем вновь фосфорилируется в УТФ за счет АТФ,
и таким образом весь цикл превращений глюкозо-1-фосфата начинается сначала.
В целом образование α-1,4-глюкозидной ветви («амилозной» ветви) гликогена можно представить в виде следующей схемы:
Глюкозо-1-фосфат |
|
|
УТФ |
|
АДФ |
Глюкозо-1-фосфат- |
|
|
-уридилилтрансфераза |
|
|
|
|
Нуклеозид- |
Пирофосфат |
дифосфат- |
|
УДФ-глюкоза |
|
киназа |
|
|
|
Гликоген |
УДФ |
АТФ |
(С6Н10О5)n
Гликоген-синтаза
Гликоген
(С6Н10О5)n+1
323
Установлено, что гликогенсинтаза неспособна катализировать образование α-(1–>6)-связи, имеющейся в точках ветвления гликогена. Этот процесс катализирует специальный фермент, получивший название гликогенветвящего фермента, или амило-(1–>4)–>(1–>6)-трансглюкозидазы. Последний катализирует перенос концевого олигосахаридного фрагмента, состоящего из 6 или 7 остатков глюкозы, с нередуцирующего конца одной из боковых цепей, насчитывающей не менее 11 остатков, на 6-гидроксиль- ную группу остатка глюкозы той же или другой цепи гликогена. В результате образуется новая боковая цепь.
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы.
Таким образом, ветвление увеличивает скорость синтеза и расщепления гликогена.
Благодаря способности к отложению гликогена (главным образом в печени и мышцах и в меньшей степени в других органах и тканях) создаются условия для накопления в норме некоторого резерва углеводов. При повышении энерготрат в организме в результате возбуждения ЦНС обычно происходят усиление распада гликогена и образование глюкозы.
Помимо непосредственной передачи нервных импульсов к эффекторным органам и тканям, при возбуждении ЦНС повышаются функции ряда желез внутренней секреции (мозговое вещество надпочечников, щитовидная железа, гипофиз и др.), гормоны которых активируют распад гликогена, прежде всего в печени и мышцах (см. главу 8).
Как отмечалось, эффект катехоламинов в значительной мере опосредован действием цАМФ, который активирует протеинкиназы тканей. При участии последних происходит фосфорилирование ряда белков, в том числе гликогенсинтазы и фосфорилазы b – ферментов, участвующих в обмене углеводов. Фосфорилированный фермент гликогенсинтаза сам по себе малоактивен или полностью неактивен, но в значительной мере активируется положительным модулятором глюкозо-6-фосфатом, который увеличивает Vmax фермента. Эта форма гликогенсинтазы называется D-формой, или зависимой (dependent) формой, поскольку ее активность зависит от глюкозо-6-фосфата. Дефосфорилированная форма гликогенсинтазы, называемая также I-формой, или независимой (independent) формой, активна и в отсутствие глюкозо-6-фосфата.
Таким образом, адреналин оказывает двойное действие на обмен углеводов: ингибирует синтез гликогена из УДФ-глюкозы, поскольку для проявления максимальной активности D-формы гликогенсинтазы нужны очень высокие концентрации глюкозо-6-фосфата, и ускоряет распад гликогена, так как способствует образованию активной фосфорилазы а. В целом суммарный результат действия адреналина состоит в ускорении превращения гликогена в глюкозу.
Распад гликогена (гликогенолиз)
Известно, что фосфоролитический распад играет ключевую роль в мобилизации полисахаридов *.
* В тканях человека и животных отечественными биохимиками Е.Л. Розенфельд и И.А. Поповой обнаружен также фермент α-амилаза, катализирующий отщепление остатков глюкозы от молекулы гликогена по α-1,4-связи. Однако ведущая роль в расщеплении гликогена в клетках принадлежит фосфорилазам.
324
Аденилатциклаза |
|
(неактивная) |
|
Адреналин, глюкагон |
|
Аденилатциклаза |
|
(активная) |
|
АТФ |
цАМФ |
Н4Р2О7 |
|
цАМФ-зависимая |
цАМФ-зависимая |
протеинкиназа |
протеинкиназа |
(активная) |
(неактивная) |
Киназа фосфорилазы b (неактивная)
|
|
+ АТФ |
|
|
|
+ Мg2+ |
|
Киназа фосфорилазы b |
|
Гликоген |
|
|
(С6Н10О5)n |
||
|
(активная) |
|
|
|
|
|
Н3РO4 |
Фосфорилаза b |
|
Фосфорилаза а |
|
(неактивная) |
4АТФ 4АДФ |
(активная) |
Глюкозо-1 |
|
|
||
|
|
|
Фосфат |
4Н3РO4 |
|
4Н2O |
Гликоген |
|
(C6H10O5)n-1 |
||
|
Фосфатаза |
|
|
|
|
|
|
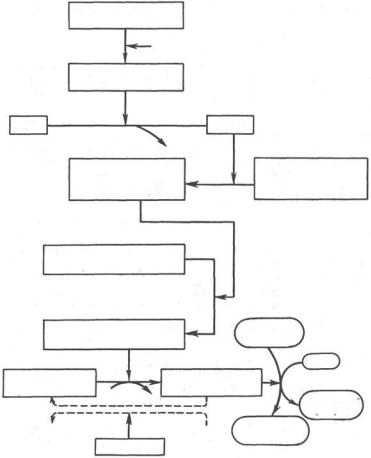
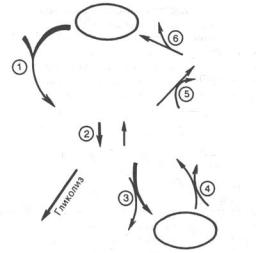
Рис. 10.1. Гормональная регуляция фосфоролитического отщепления остатка глюкозы от гликогена.
Фосфорилазы переводят полисахариды (в частности, гликоген) из запасной формы в метаболически активную форму; в присутствии фосфорилазы гликоген распадается с образованием фосфорного эфира глюкозы (глюкозо-1-фосфата) без предварительного расщепления на более крупные обломки молекулы полисахарида. В общей форме эту реакцию можно представить в следующем виде:
(C6H10O5)n + H3PO4 –> (C6H10O5)n–1 + Глюкозо-1-фосфат,
где (С6Н10О5)n означает полисахаридную цепь гликогена, а (С6Н10О5)n,– ту же цепь, но укороченную на один глюкозный остаток.
На рис. 10.1 изображены процесс распада гликогена до глюкозо-1-фосфата и участие в этом процессе цАМФ. Фермент фосфорилаза существует в двух формах, одна из которых (фосфорилаза а) активна, в то время как другая (фосфорилаза b) обычно неактивна. Обе формы могут диссоциировать на субъединицы. Фосфорилаза b состоит из двух субъединиц, а фосфорилаза а–из четырех. Превращение фосфорилазы b в фосфорилазу а осуществляется фосфорилированием белка:
325
2 Фосфорилаза b + 4 АТФ –> Фосфорилаза а + 4 АДФ.
Катализируется эта реакция ферментом, который называется киназой фосфорилазы b. Установлено, что эта киназа может существовать как в активной, так и в неактивной форме. Неактивная киназа фосфорилазы превращается в активную под влиянием фермента протеинкиназы (киназа киназы фосфорилазы), и не просто протеинкиназы, а цАМФ-зависимой протеинкиназы.
Активная форма последней образуется при участии цАМФ, которая в свою очередь образуется из АТФ под действием фермента аденилатциклазы, стимулируемой, в частности, адреналином и глюкагоном. Увеличение содержания адреналина в крови приводит в этой сложной цепи реакций к превращению фосфорилазы b в фосфорилазу а и, следовательно, к освобождению глюкозы в виде глюкозо-1-фосфата из запасного полисахарида гликогена. Обратное превращение фосфорилазы а в фосфорилазу b катализируется ферментом фосфатазой (эта реакция практически необратима).
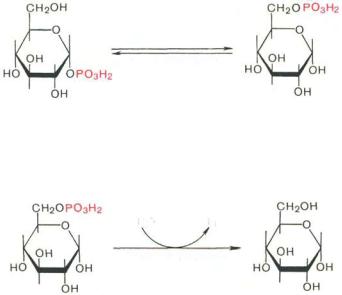
Образовавшийся в результате фосфоролитического распада гликогена глюкозо-1-фосфат превращается под действием фосфоглюкомутазы в глюкозо-6-фосфат. Для осуществления данной реакции необходима фосфорилированная форма фосфоглюкомутазы, т.е. ее активная форма, которая образуется, как отмечалось, в присутствии глюкозо-1,6-бисфосфата *.
Фосфоглюкомутаза
Глюкозо-1-фосфат |
Глюкозо-6-фосфат |
|
Образование свободной глюкозы из глюкозо-6-фосфата в печени происходит под влиянием глюкозо-6-фосфатазы. Данный фермент катализирует гидролитическое отщепление фосфата:
Н2O Pi
Глюкозо-6-фосфатаза
Глюкозо-6-фосфат |
Глюкоза |
* В настоящее время установлено, что в каталитическом центре активной формы молекулы фосфоглюкомутазы присутствует фосфорилированный остаток серина. Во время катализа эта фосфорильная группа, вероятно, переносится на гидроксильную группу при С-6 глюкозо-1-фосфата с образованием глюкозо-1-бисфосфата. Далее фосфорильная группа указанного промежуточного продукта переносится на остаток серина в активном центре. В результате происходят образование глюкозо-6-фосфата и регенерирование фосфорилированного фермента.
326
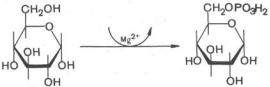
|
Гликоген |
УДФ |
|
|
|
||
|
Н3РО4 |
|
|
|
|
УДФ-глюкоза |
|
|
|
Пирофосфат |
|
|
Глюкозо-1-фосфат |
УТФ |
|
|
|
||
Рис. 10.2. Распад и синтез гликогена |
Глюкозо-6-фосфат |
|
|
(схема). |
АДФ |
||
|
|||
Жирными стрелками указан путь распада, |
Н2О |
|
|
тонкими - путь синтеза. Цифрами обозна- |
|
|
|
чены ферменты: 1 - фосфорилаза; 2 - фос- |
|
АТФ |
|
фоглюкомутаза; 3 - глюкозо-6-фосфатаза; |
|
||
|
|
||
4 - гексокиназа (глюкокиназа); 5 - глюко- |
|
Глюкоза |
|
зо-1-фосфат-уридилтрансфераза; 6- глико- |
|
||
Н3РО4 |
|
||
генсинтаза. |
|
||
|
|
Заметим, что фосфорилированная глюкоза в противоположность неэтерифицированной глюкозе не может легко диффундировать из клеток. Печень содержит гидролитический фермент глюкозо-6-фосфатазу, который
иобеспечивает возможность быстрого выхода глюкозы из этого органа.
Вмышечной ткани глюкозо-6-фосфатаза практически отсутствует.
На рис. 10.2 отражены представления о путях распада и синтеза гликогена в печени *.
Можно считать, что сохранение постоянства концентрации глюкозы в крови является результатом одновременного протекания двух процессов: поступления глюкозы в кровь из печени и потребления ее из крови тканями, где она используется в первую очередь как энергетический материал.
В тканях (в том числе в печени) распад глюкозы происходит двумя основными путями: анаэробным (при отсутствии кислорода) и аэробным, для осуществления которого необходим кислород.
ГЛИКОЛИЗ
Гликолиз (от греч. glycys–сладкий и lysis–растворение, распад)–это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ.
При аэробных условиях пируват проникает в митохондрии, где полностью окисляется до СО2 и Н2О. Если содержание кислорода недостаточно, как это может иметь место в активно сокращающейся мышце, пируват превращается в лактат.
Итак, гликолиз–не только главный путь утилизации глюкозы в клетках, но и уникальный путь, поскольку он может использовать кислород, если
* Как отмечалось, в отличие от печени в мышечной ткани глюкозо-6-фосфатаза отсутствует. Пути распада и синтеза гликогена в печени в целом подобны таковым в мышце, однако имеются существенные различия в структуре печеночных и мышечных ферментов метаболизма, а также в механизмах регуляции их активности.
327
последний доступен (аэробные условия), но может протекать и в отсутствие кислорода (анаэробные условия).
Анаэробный гликолиз –сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:
С6Н12О6 + 2АДФ + 2ФН –> 2СН3СН(ОН)СООН + 2АТФ + 2Н2О.
Глюкоза Молочная кислота
В анаэробных условиях гликолиз–единственный процесс в животном организме, поставляющий энергию. Именно благодаря гликолизу организм человека и животных определенный период может осуществлять ряд физиологических функций в условиях недостаточности кислорода. В тех случаях, когда гликолиз протекает в присутствии кислорода, говорят об
аэробном гликолизе *.
Последовательность реакций анаэробного гликолиза, так же как и их промежуточные продукты, хорошо изучена. Процесс гликолиза катализируется одиннадцатью ферментами, большинство из которых выделено в гомогенном, клисталлическом или высокоочищенном виде и свойства которых достаточно известны. Заметим, что гликолиз протекает в гиалоплазме (цитозоле) клетки.
Первой ферментативной реакцией гликолиза является фосфорилирование, т.е. перенос остатка ортофосфата на глюкозу за счет АТФ. Реакция катализируется ферментом гексокиназой:
АТФ |
АДФ |
Гексокиназа |
|
Глюкоза |
Глюкозо-6-фосфат |
Образование глюкозо-6-фосфата в гексокиназной реакции сопровождается освобождением значительного количества свободной энергии системы и может считаться практически необратимым процессом.
Наиболее важным свойством гексокиназы является ее ингибирование глюкозо-6-фосфатом, т.е. последний служит одновременно и продуктом реакции, и аллостерическим ингибитором.
Фермент гексокиназа способен катализировать фосфорилирование не только D-глюкозы, но и других гексоз, в частности D-фруктозы, D-маннозы и т.д. В печени, кроме гексокиназы, существует фермент глюкокиназа, который катализирует фосфорилирование только D-глюкозы. В мышечной ткани этот фермент отсутствует (подробнее см. главу 16).
* В аэробных условиях гликолитический распад глюкозы до пировиноградной кислоты можно рассматривать как первую стадию окисления глюкозы до конечных продуктов этого
процесса - СО2 и Н2О.
328
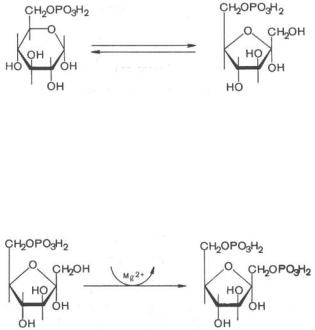
Второй реакцией гликолиза является превращение глюкозо-6-фос- фата под действием фермента глюкозо-6-фосфат-изомеразы во фруктозо-6- фосфат:
Глюкозо-6-фосфат- изомераза
Глюкозо-6-фосфат |
Фруктозо-6-фосфат |
Эта реакция протекает легко в обоих направлениях, и для нее не требуется каких-либо кофакторов.
Третья р е а к ц и я катализируется ферментом фосфофруктокиназой; образовавшийся фруктозо-6-фосфат вновь фосфорилируется за счет второй молекулы АТФ:
АТФ |
АДФ |
6-Фосфофруктокиназа |
|
Фруктозо-6-фосфат |
Фруктозо-1,6-бисфосфат |
Данная реакция аналогично гексокиназной практически необратима, протекает в присутствии ионов магния и является наиболее медленно текущей реакцией гликолиза. Фактически эта реакция определяет скорость гликолиза в целом.
Фосфофруктокиназа относится к числу аллостерических ферментов. Она ингибируется АТФ и стимулируется АМФ *. При значительных величинах отношения АТФ/АМФ активность фосфофруктокиназы угнетается и гликолиз замедляется. Напротив, при снижении этого коэффициента интенсивность гликолиза повышается. Так, в неработающей мышце активность фосфофруктокиназы низкая, а концентрация АТФ относительно высокая. Во время работы мышцы происходит интенсивное потребление АТФ и активность фосфофруктокиназы повышается, что приводит к усилению процесса гликолиза.
Четвертую реакцию гликолиза катализирует фермент альдолаза. Под влиянием этого фермента фруктозо-1,6-бисфосфат расщепляется на две фосфотриозы:
* Активность фосфофруктокиназы ингибируется также цитратом. Показано, что при диабете, голодании и некоторых других состояниях, когда интенсивно используются жиры как источник энергии, в клетках тканей содержание цитрата может возрастать в несколько раз. В этих условиях происходит резкое торможение активности фосфофруктокиназы цитратом.
329
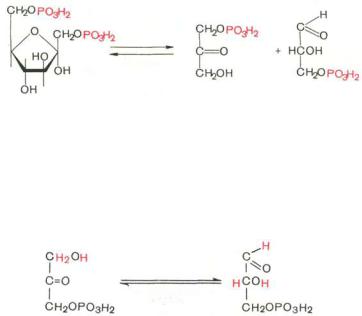
|
Альдолаза |
|
|
Диоксиацетон- |
Глицеральде- |
Фруктозо-1,6-бисфосфат |
фосфат |
гид-3-фосфат |
Эта реакция обратима. В зависимости от температуры равновесие устанавливается на различном уровне. При повышении температуры реакция сдвигается в сторону большего образования триозофосфатов (дигидроксиацетонфосфата и глицеральдегид-3-фосфата)*.
Пятая р е а к ц и я–это реакция изомеризации триозофосфатов. Катализируется ферментом триозофосфатизомеразой:
Триозофосфатизомераза
Диоксиацетон- |
Глицеральдегид- |
фосфат |
3-фосфат |
Равновесие данной изомеразной реакции сдвинуто в сторону дигидроксиацетонфосфата: 95% дигидроксиацетонфосфата и около 5% глице- ральдегид-3-фосфата. В последующие реакции гликолиза может непосредственно включаться только один из двух образующихся триозофосфатов,
аименно глицеральдегид-3-фосфат. Вследствие этого по мере потребления
входе дальнейших превращений альдегидной формы фосфотриозы дигидроксиацетонфосфат превращается в глицеральдегид-3-фосфат.
Образованием глицеральдегид-3-фосфата как бы завершается первая
стадия гликолиза. Вторая стадия–наиболее сложная и важная. Она включает окислительно-восстановительную реакцию (реакция гликолитической оксидоредукции), сопряженную с субстратным фосфорилированием, в процессе которого образуется АТФ.
В результате шестой реакции глицеральдегид-3-фосфат в присутствии фермента глицеральдегидфосфатдегидрогеназы, кофермента НАД и неорганического фосфата подвергается своеобразному окислению с образованием 1,3-бисфосфоглицериновой кислоты ** и восстановленной формы НАД (НАДН). Эта реакция блокируется йодили бромацетатом, протекает
внесколько этапов:
*Животные ткани содержат по меньшей мере три различные альдолазы, характерные для мышцы, печени и мозга соответственно. Все альдолазы расщепляют фруктозо-1,6-бисфосфат до диоксиацетонфосфата и глицеральдегид-3-фосфата и могут катализировать обратную конденсацию диоксиацетонфосфата с различными оксиальдегидами, хотя и с неодинаковой
скоростью.
** Глицеральдегид-3-фосфат - последний углевод в цепи превращений глюкозы. Дальнейшим превращениям подвергаются органические кислоты, которые находятся в диссоциированной форме, поэтому наряду с названием свободных кислот используют также название их анионов, например 3-фосфоглицерат, пируват и др.
330