

tema_1_fiziologia_myshechnykh_kletok_1
.docx
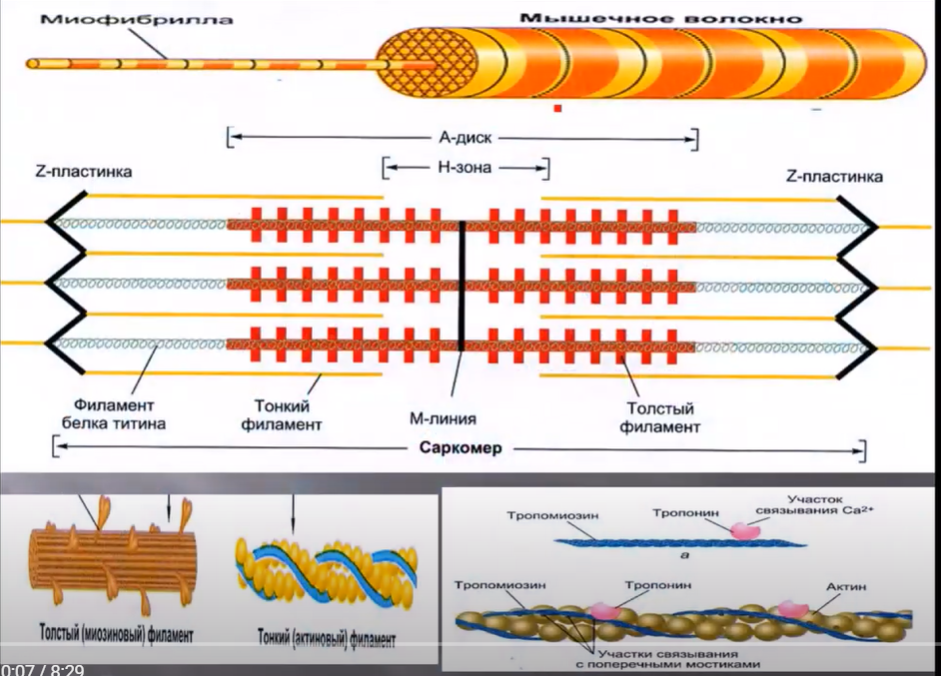
У человека более 600 скелетных мышц, однако механизм сокращения этих мышц одинаков. Сокращение скелетных мышц обусловлено укорочением мышечного волокна. В свою очередь укорочение мышечного волокна связано с укорочением саркомера. Саркомер ограничен Z-пластинками. Эти пластинки состоят из переплетенных белков, в том числе белка десмина. Появление данного белка в саркоплазме вне Z-пластинки является признаком миастении. Сам саркомер представлен двумя филаментами – это тонкие филаменты, которые состоят из актиновых нитей и толстые филаменты, которые состоят из миозиновых нитей. И актиновые и миозиновые нити формируются сократительными белками. В свою очередь у актиновых нитей имеются центры связывания с головкой поперечного мостика миозина. Именно этот процесс связывания способствует укорочению саркомера.
Тонкие филаменты, т.е. актиновые нити взаимодействуют с Z-пластинками, т.е. прикреплены к этим пластинкам. Толстые филаменты прикреплены к М-линии. М-линия, также как и Z-пластинки, образованы белками, и за счет белка титина объединяют миозиновые нити и их центры с Z-пластинкой. Кроме того, титин обладает упругими свойствами и способствует упруго растяжимым свойствам саркомера.
Саркомер представлен несколькими дискамии зонами. А-диск – в состав этого диска входит как тонкий, так и толстый филамент, т.е. актиновые и миозиновые нити. Н-зона представлена только миозиновыми нитями. I-диск представлен только актиновыми нитями.
В состав саркомера также входят регуляторные белки тропонин и тропомиозин. Тропонин представлен 3 субъединицами и связан с тропомиозином. Кроме того, у тропонина имеется участок связывания с кальцием. Сам тропомиозин блокирует активные центры актина, препятствуя взаимодействию головки поперечного мостика с данными центрами. Но для такой блокады необходимо наличие связи с тропонином.
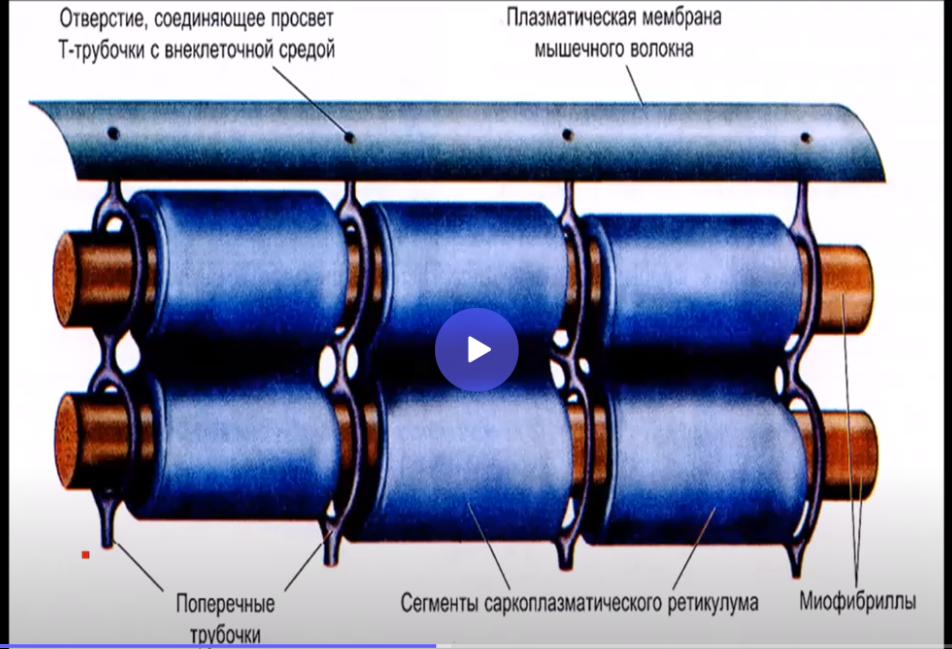
Для понимания сокращения скелетных мышц, а именно электромеханического сопряжения, необходимо еще знать, что в скелетных мышцах, а точнее в мышечных волокнах имеются Т-трубочки, которые представляют собой впячивания сарколеммы. Эти Т-трубочки через дигидропиридиновые рецепторы реагируют на изменение мембранного потенциала и активируют рианодиновые рецепторы саркоплазматического ретикулума, т.е. ЭПС, где находятся ионы кальция.
В основе сокращения скелетных мышц лежит теория скользящих нитей. Эта теория базируется на электромеханическом сопряжении, которое в свою очередь состоит из 2 частей: электрохимического и хемомеханического. Все начинается с выделения медиатора. В нервно-мышечном синапсе данным медиатором является ацетилхолин. Взаимодействие этого медиатора с концевой пластинкой мышечного волокна, т.е. с постсинаптической мембраной, приводит в начале к развитию местной деполяризации, а точнее потенциала концевой пластинки, и при достижении критического уровня развивается потенциал действия. Этот ПД распространяется вдоль мембраны мышечного волокна с вовлечением в процесс Т-трубочек. От этих впячиваний сарколеммы через дигидропиридиновые рецепторы передается возбуждение к рианодиновым рецепторам саркоплазматического ретикулума. Это является пусковым стимулом для выхода ионов кальция в саркоплазму. На этом электрохимическое сопряжение заканчивается и начинается хемомеханическое сопряжение. Взаимодействие ионов кальция с регуляторным белком тропонином приводит к изменению конфигурации этого белка и как следствие к разрыву связей тропонина с тропомиозином. В результате открываются активные центры актина. Это притягивает головку миозина к активному центру актина. При таком взаимодействии поперечный мостик миозина приобретает АТФ-активность и тем самым происходит расщепление АТФ с выделением энергии и с последующим отсоединением головки миозина. Таким образом, при взаимодействии АТФ с поперечным мостиком миозина, с последующим вращением головки и отсоединением, происходит скольжение актина и миозина по отношению друг к другу, что приводит к укорочению саркомера, и как следствие укорочению мышечного волокна и сокращению мышцы.