

3.5.2.3. Морфология хромосом
Митотическая суперкомпактизация хроматина делает возможным изучение внешнего вида хромосом с помощью световой микроскопии. В первой половине митоза они состоят из двух хроматид, соединенных между собой в области первичной перетяжки (центромеры или кинетохора) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы, распределяющиеся между дочерними клетками.
В зависимости от места положения центромеры и длины плеч, расположенных по обе стороны от нее, различают несколько форм хромосом: равноплечие, или метацентрические (с центромерой посередине), неравноплечие, или субметацентрические (с центромерой, сдвинутой к одному из концов), палочковидные, или акроцентрические (с центромерой, расположенной практически на конце хромосомы), и точковые —очень небольшие, форму которых трудно определить (рис. 3.52). При рутинных методах окраски хромосом они различаются по форме и соотносительным размерам. При использовании методик дифференциальной окраски выявляется неодинаковая флуоресценция или распределение красителя по длине хромосомы, строго специфические для каждой отдельной хромосомы и ее гомолога (рис. 3.53).
Таким образом, каждая хромосома индивидуальна не только по заключенному в ней набору генов, но и по морфологии и характеру дифференциального окрашивания.
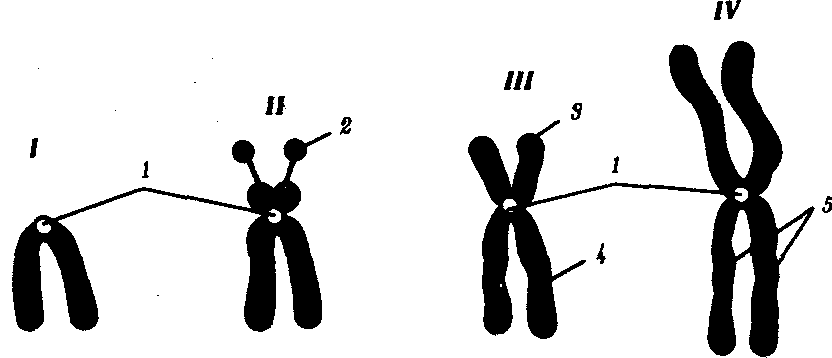
Рис. 3.52. Формы хромосом:
I — телоцентрическая, II — акроцентрическая, III—субметацентрическая, IV—метацентрическая;
1 — центромера, 2 — спутник, 3 — короткое плечо, 4 — длинное плечо, 5 — хроматиды
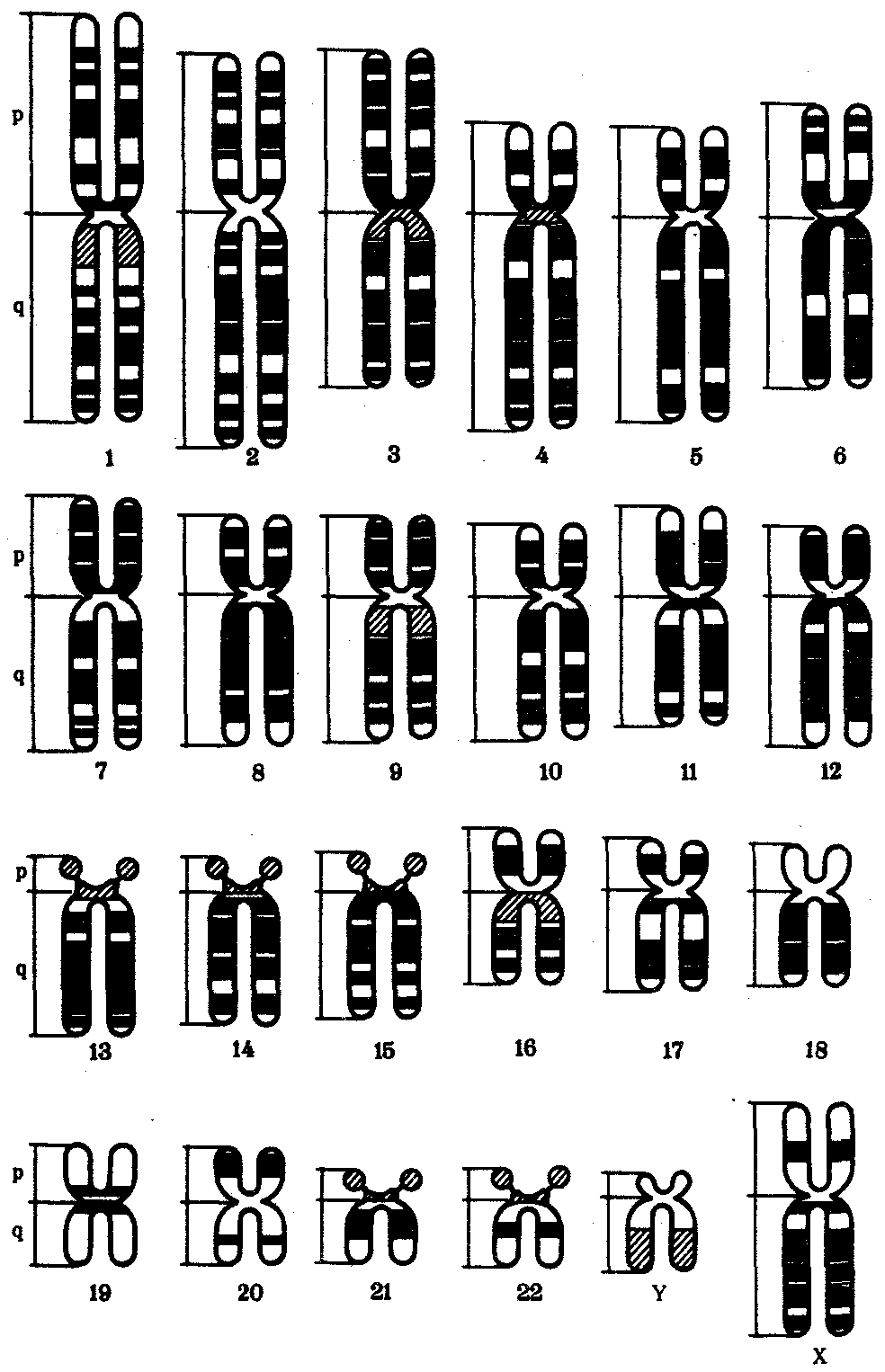
Рис. 3.53. Расположение локусов в хромосомах человека
при их дифференциальном окрашивании:
р — короткое плечо, q — длинное плечо; 1—22 — порядковый номер хромосомы; XY — половые хромосомы
3.5.2.4. Особенности пространственной организации генетического материала в прокариотической клетке
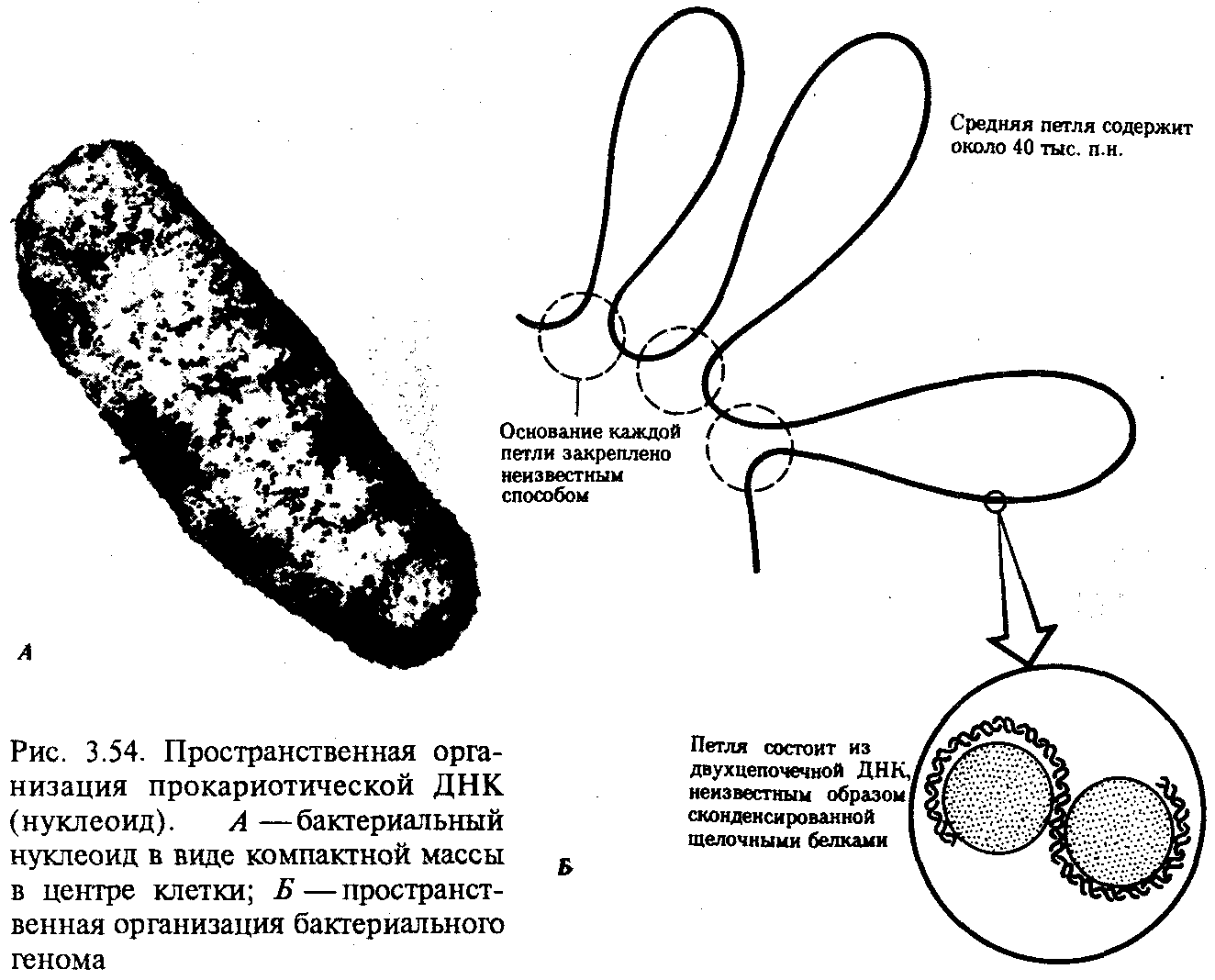
Описанная организация наследственного материала эукариот, связанная с расположением его функциональных единиц в ядерных структурах—хромосомах, у прокариот отсутствует. Основная часть генетического материала заключена у них в единственной кольцевой молекуле ДНК длиной около 1 мм (Е. coli), причем у прокариот не обнаружено гистонов, обеспечивающих нуклеосомную организацию хроматина эукариот. Однако кольцевой молекуле ДНК прокариот также присуща укладка в виде петель. ДНК прокариот тоже комплексируется с некоторыми негастоновыми белками, образуя нуклеоид, который гораздо менее устойчив по сравнению с хроматином эукариот (рис. 3.54).
3.5.3. Проявление основных свойств материала наследственности и изменчивости на хромосомном уровне его организации
На хромосомном уровне организации, который появляется в процессе эволюции у эукариотических клеток, генетический аппарат должен удовлетворять всем требованиям, предъявляемым к субстрату наследственности и изменчивости: обладать способностью к самовоспроизведению, поддержанию постоянства своей организации и приобретению изменений, которые могут передаваться новому поколению клеток.
3.5.3.1. Самовоспроизведение хромосом в митотическом цикле клеток

Рис. 3.55. Возникновение двунитчатой хромосомы из однонитчатой:
I — интерфазная хромосома до репликации ДНК,
II — интерфазная хромосома после репликации ДНК,
III — метафазная хромосома
Процесс самовоспроизведения — редупликации хромосом осуществляется в период подготовки клетки к митозу — в интерфазе. Вследствие этого в ходе после- дующего деления клетка передает дочернему поколению полноценный комплекс генов, заключенный в каждой отдельной хромосоме. В основе самовоспроизведения хромосомы лежит репликация ДНК с образованием из одной материнской спирали двух дочерних молекул, на базе которых образуются две сестринские хроматиды (рис. 3.55). Благодаря высокой точности репликации ДНК (см. разд. 3.4.2.2 ) сестринские хроматиды несут практически одинаковую информацию, записанную в ее дочерних молекулах. В каждой хроматиде ДНК находится в комплексе с гистонами, и ей присущи все описанные выше уровни компактизации хроматина, свойственные интерфазной клетке.
Таким образом, готовясь к самовоспроизведению, клетка удваивает содержание ДНК в каждой хромосоме. При этом последняя приобретает двунитчатую структуру.
Известны случаи, когда репликация ДНК, причем неоднократная, не сопровождается формированием хроматид на базе дочерних спиралей. При этом образуются так называемые политенные хромосомы, содержащие многие сотни копий ДНК. Такие хромосомы обнаруживаются, например, в неделящихся клетках слюнных желез личинок некоторых насекомых при обычной световой микроскопии (рис. 3.56).
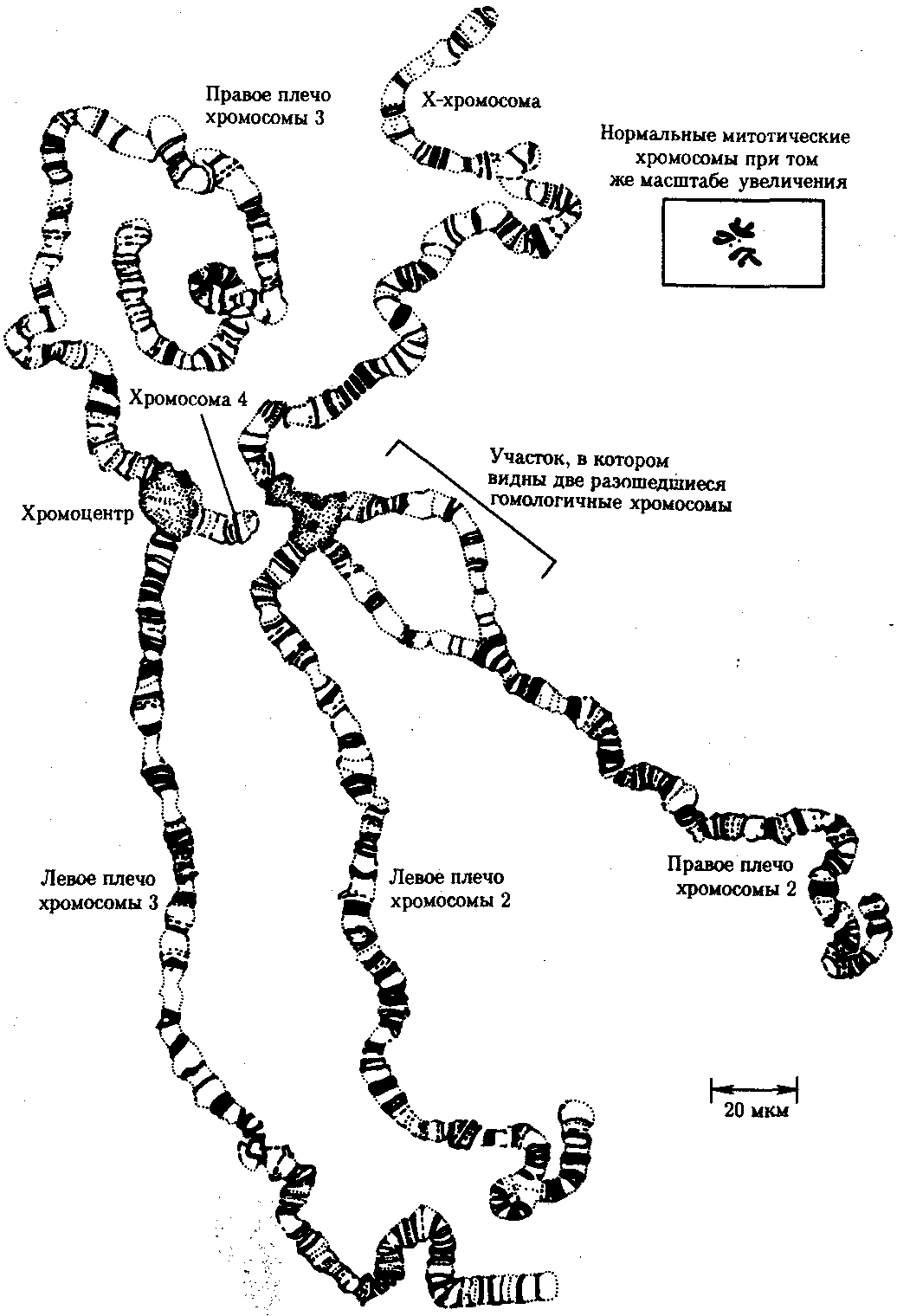
Рис. 3.56. Полный набор политенных хромосом
из клетки слюнной железы дрозофилы