Механизм световосприятия
Рецептор, воспринимающий фотон света – это молекула, состоящая из:
опсина (белок ) + ретиналя (альдегид витамина А).
Поглощение родопсином кванта света вызывает ряд его химических превращений – фотолиз. При попадании фотона на родопсин: Цис-ретиналь → транс-ретиналь, рецепторная молекула распадается на составляющие, а в темноте происходит ее ресинтез
Ретиналь везде одинаковый, отличаются белки-опсины.
В палочках пигмент родопсин, который распадается при попадании на него волны любой длины, поэтому палочки не способны различать цвета (сумеречное зрение)
 В
колбочках несколько видов йодопсинов:
В
колбочках несколько видов йодопсинов:
хлоролаб («зеленый»),
эритролаб («красный»)
цианолаб («синий»).
Каждый из них начинает распадаться при попадании на него световой волны только
определённой длины (от 390 до 760 нм). Поэтому колбочки могут различать цвета.
В темноте часть воротных каналов находится в открытом состоянии, и ионы Na+ и Са2+ могут свободно диффундировать из внеклеточного пространства в цитозоль. Поток ионов в темноте, или темновой ток (был открыт в 1970 году Вильямом Хейгинсом), вызывает деполяризацию плазматической мембраны наружного сегмента палочки. В темноте потенциал мембраны наружного сегмента составляет около -50 или -40 мВ вместо - 70 мВ для нормальной нервной клетки. То есть в темноте фоторецепторные клетки более деполяризованы, чем обычные нервные клетки в состоянии покоя, а деполяризация вызывает непрерывное высвобождение медиатора из окончаний их аксонов – в точности так, как это происходит в обычных рецепторах при стимуляции.
В результате поглощения кванта света молекулой родопсина или йодопсина и следующих за этим биохимических реакций происходит закрытие ионных каналов (Na+/Са2+), что приводит к уменьшению темнового тока и гиперполяризации (увеличению наружного положительного заряда) плазматической мембраны клетки. Свет, повышая потенциал на мембране рецепторной клетки (гиперполяризуя ее), уменьшает выделение медиатора. Пполучается, что стимуляция выключает рецепторы и приостанавливает выделение медиатора в синаптическую щель. .
Процесс кодирования световой информации родопсином палочки

В результате описанных процессов каскад ферментативных реакций, приводящий к временной остановке выделения медиатора можно представить в следующей последовательности:
1. Молекула родопсина поглощает 1 фотон > распадается на трансретиналь и опсин
2. Активируется около 500 молекул G-белка>
5. Закрываются около 250 натриевых каналов >
6. Уменьшается поступления ионов Na+ в клетку на 106-107 ионов >
7. Мембрана палочки гиперполяризуется до 1мВ.
Т.о. рецепторный потенциал фоторецепторной клетки - гиперполяризационный, связанный с закрытием каналов и прекращением тока натрия (больше) и кальция (меньше) в клетку. Он электротонический и градуированный - его величина зависит от количества воспринятых фотонов.
На свету родопсин распадается, в темноте ресинтезируется (поэтому интуитивно закрываем уставшие глаза)
· При равномерном постоянном освещении: равновесие:
скорость распада родопсина = скорости его ресинтеза.
· При уменьшении освещенности равновесие смещается:
скорость распада < скорости ресинтеза
Т.к. родопсина стало больше → больше чувствительность к свету
Это темновая адаптация –происх. за 30-60 мин:
- через 1 мин – чувствительность к свету увелич. в 10раз
- через 20 мин – в 6 000 раз
- через 40 мин – в 25 000 раз
Нарушение темновой адаптации – «куриная слепота» (ночная слепота)!!!
· При увеличении освещенности:
скорость распада > скорости ресинтеза → меньше чувствительность к свету (охранительная функция). Это световая адаптация. Осуществляется за 60 с
Темновая адаптация – процесс приспособления зрения к измененным условиям освещенности (к недостатку освещенности), при котором увеличивается фоточувствительность глаза. Адаптация (лат. adapto – приспособляю) глаза человека и животных огромна
Адаптация осуществляется все время, пока человек находится в темноте. Она происходит без контроля сознания. Сначала процесс адаптации идет более быстро, затем замедляется. Особенно резко повышается чувствительность в первые полчаса пребывания в темноте. Световая чувствительность при темновой адаптации достигает своего максимального значения примерно через 60-80 мин. после начала адаптации, при этом время адаптации зависит от того, к какой освещенности был адаптирован глаз перед понижением освещенности (чем больше была освещенность, тем дольше идет адаптация). Также скорость адаптации зависит от возраста. Наибольшая скорость адаптации у людей в возрасте 20-25 лет, а в пожилом возрасте она уменьшается в 8-10 раз, по отношению к максимальному значению.
При темновой адаптации происходит ряд изменений в зрительной сенсорной системе.
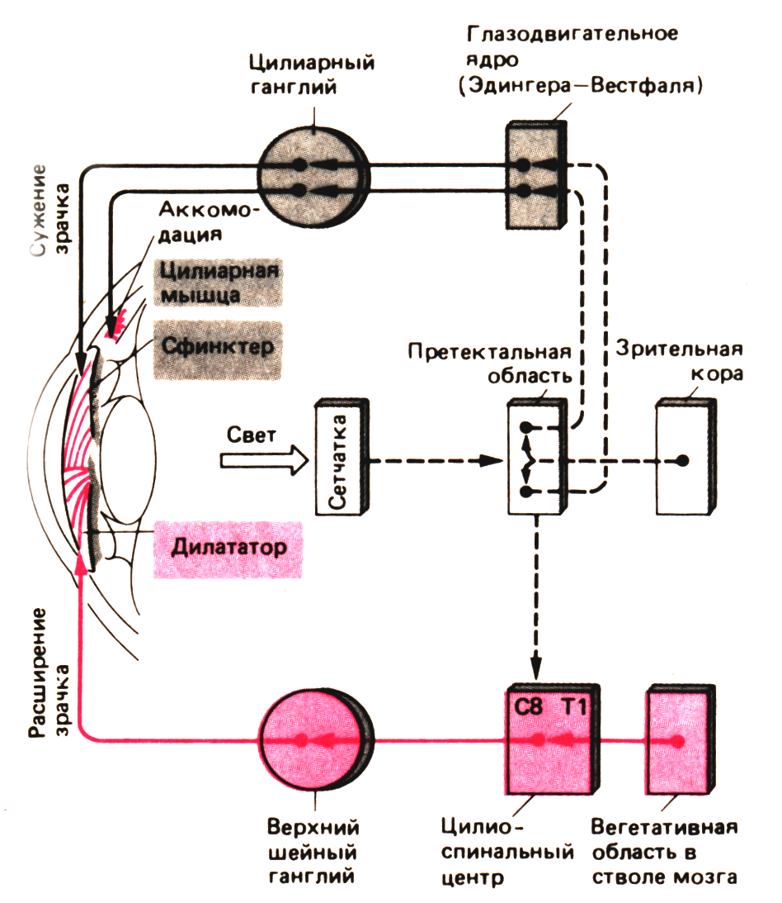
Во-первых, изменение диаметра зрачка. Если учесть, что диаметр зрачка при ярком освещении равен 2 мм, то у некоторых молодых людей зрачок в темноте расширяется до 8 мм, то есть увеличивается в 4 раза. Это означает, что глаз может собрать в 16 раз больше света. Но изменение диаметра зрачка – не основополагающий процесс в темновой адаптации. Расширение зрачка происходит медленнее, чем сужение. Так, после выключения света за первые 3-4 с зрачок расширяется только до 6 мм, еще за 40 с он расширяется до 7 мм и лишь за 3 мин. достигает своего максимального диаметра. Характерно, но с возрастом максимальный диаметр зрачка уменьшается.
Во-вторых, одновременно с расширением зрачка нарастает концентрация светочувствительных пигментов (родопсина и йодопсина) в фоторецепторных клетках сетчатки. Возрастание концентрации пигментов происходит медленно (на протяжении 10 мин. и более), но оно во много раз увеличивает светочувствительность. Нарастание концентрации происходит за счет восстановления данныз пигментов.
В-третьих, переход с колбочкового зрения на палочковое. При высокой освещенности функционируют колбочки, и они способны обеспечивать высокую остроту зрения и цветовосприятие. При низком уровне освещенности функционируют преимущественно палочки, потому что они намного чувствительнее к свету, но они не обеспечивают большую остроту зрения. Переход на палочковое зрение приводит к тому, что человек в сумерках перестает различать цвета
В-четвертых, темновая адаптация сопровождается медленной ретиномоторной реакцией. В ходе нее темные пигментные гранулы меланина, которые находятся в отростках пигментного эпителия и окружают фоторецепторные клетки сетчатки, перемещаясь из отростков в тела клеток эпителия. Тем самым они открывают доступ света к фоторецепторным клеткам. При этом опять же повышается светочувствительность палочек.
Световая адаптация – это приспособление органа зрения к условиям высокой освещенности. Она протекает очень быстро, в отличие от темновой адаптации. Слишком яркий свет вызывает неприятное ощущение ослепления, потому что раздражение палочек из-за слишком быстрого разложения родопсина чрезвычайно сильно, они «ослеплены». Даже колбочки, которые не защищены еще зернами черного пигмента меланина, раздражены слишком сильно.
Верхняя граница слепящей яркости зависит от времени темновой адаптации глаза: чем дольше была темновая адаптация, тем меньшая яркость света вызывает ослепление. Если в поле зрения попадают очень ярко освещенные (слепящие) объекты, то они ухудшают восприятие сигналов на большей части сетчатки. Только по истечении достаточного времени приспособление глаза к яркому свету заканчивается, прекращается неприятное чувство ослепления и глаз начинает нормально функционировать. Полная световая адаптация длится от 8 до10 мин.
При световой адаптации начинает работать колбочковый аппарат сетчатки. Если до этого освещение было слабое, то с палочкового зрения глаз переходит на колбочковое, зрачок сужается, все это сопровождается медленной ретиномоторной реакцией.
Рассмотрим более подробно эти механизмы приспособления глаза к яркому свету.
Во-первых, происходит сужение зрачка. Если при затемнении зрачок расширяется, то на свету он быстро сужается (зрачковый рефлекс) и регулирует поток света, попадающий в глаз. При ярком свете кольцевая мышца радужки сокращается, а радиальная расслабляется. В результате зрачок сужается и уменьшается световой поток, этот процесс предотвращает повреждение сетчатки. Так, на ярком свету диаметр зрачка уменьшается до 1.8 мм, а при средней дневной освещенности он составляет около 2.4 мм.
Во-вторых, переход с палочкового зрения на колбочковое (в пределах нескольких мс). При этом чувствительность колбочек уменьшается для восприятия большей яркости, а палочки в это время углубляются немного в слой колбочек. Этот процесс является обратным тому, что происходит при темновой адаптации.
В-третьих, выцветание родопсина ,снижение скорости его ресинтеза. На свету ретиналь превращается в витамин А, который перемещается в клетки пигментного слоя (весь этот процесс и называется выцветанием родопсина).
 В-четвертых,
в пигментном слое клеток, содержащих
черный пигмент меланин, происходит
миграция гранул пигмента в отростки,
частично перекрывающие наружные сегменты
фоторецепторов, увеличивая поглощение
пришедших через сетчатку световых
лучей.
В-четвертых,
в пигментном слое клеток, содержащих
черный пигмент меланин, происходит
миграция гранул пигмента в отростки,
частично перекрывающие наружные сегменты
фоторецепторов, увеличивая поглощение
пришедших через сетчатку световых
лучей.
Схема зрачкового рефлекса
Слой горизонтальных клеток:
· возбуждаются от фоторецепторных клеток,
· тормозят биполярные клетки, но не “своего”, а соседнего сенсорного канала - латеральное торможение
Слой биполярных клеток
Возбуждаются или тормозятся непосредственно от фоторецепторных клеток. Являются первыми чувствительными нейронами на пути проведения зрительной сенсорной информации. Однако, в отличие от других сенсорных систем, в данных биполярных клетках ПД не генерируется! На выделение или прекращение выделения медиатора, биполярные клетки отвечают формированием электротонического ответа. По реакции на раздражитель биполярные клетки делятся на:
· Биполяры on-типа - возбуждаются (деполяризация) на свету.
Причина- возбуждённый ф/р → непосредственно биполярная клетка
· Биполяры оff-типа: тормозятся (гиперполяризация) на свету.
Причина- возбуждённый ф/р → горизонтальная клетка (тормозная) → биполярная клетка. Это облегчает контрастирование, различение границ предметов.
Степень конвергенции фоторецепторов на биполярной клетке:
- в центре сетчатки: одна б/кл. получает сигнал только от одной колбочки – это обеспечивает максимальную остроту зрения в жёлтом пятне;
- на периферии: на одну б/кл. конвергируют до 100 палочек – такая пространственная суммация помогает видеть в темноте
Слой амакриновых клеток.
· тормозные нейроны (Медиаторы - ГАМК, глицин, дофамин, ацетилхолин, индоламин)
· возбуждаются от ганглиозных,
· тормозят ганглиозные клетки, но соседней цепочки (латеральное торможение)
Значение латерального торможения (как амакриновыми, так и ганглиозными клетками ):
1 - вычленяется только одна линия передачи возбуждения. При этом происходят потери в количестве информации, но зато достигается контрастность изображения за счёт отсечения «лишней» информации.
2- формируются центр и периферия рецепторного поля
Слой ганглиозных клеток (ГК)
Это второй нейрон по ходу проведения зрительной сенсорной информации. И именно в нем впервые генерируется ПД по принципу "всё или ничего" и передаётся далее по зрительному нерву на значительные расстояния. Ганглиозные клетки:
· Получают импульсы от биполярных клеток через трансмиттер
· Имеют фоновую активность – 5 импульсов в сек в полной темноте
· Аксоны ганглиозных клеток формируют зрительный нерв, который прободает сетчатку – слепое пятно (нет ни фоторецепторов, ни других слоев клеток)
Рецептивное поле ГК – это совокупность фоторецепторов, посылающих сигналы к одной ГК. В среднем, на 1 ганглиозную клетку конвергируют импульсы от 60 палочек и 2 колбочек.
Формирование разных сенсорных каналов:
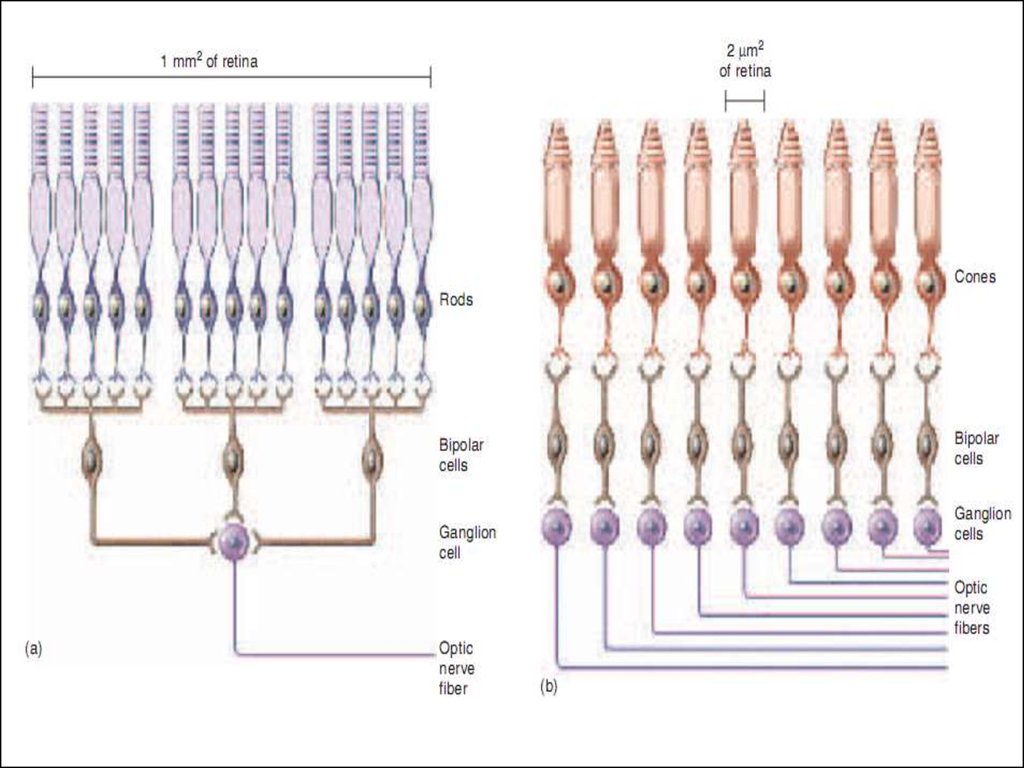
1. на периферии (рисунок а)– конвергенция по принципу сужающейся воронки, что даёт возможность суммироваться слабым сигналам при слабой освещённости: 50 фоторецепторных клеток → 1 биполярная клетка ; 4 биполярных клетки → 1 ганглиозная клетка.
В итоге: рецептивное поле 1 ганглиозной клетки- 200 фоторецепторных клеток.
2. в центре (рисунок b)– по принципу “точка-в-точку", что обеспечивает чёткое видение и высокую остроту зрения: 1 фоторецепторная клетка (колбочка) → 1 биполярная клетка → 1 ганглиозная клетка
В итоге: рецептивное поле 1 гангл/клетки: 1 колбочка
Ганглиозные клетки по характеру возбуждения рецептивного поля делятся на:
ганглиозные клетки с on- рецептивным полем:
1) при освещении центра рецептивного поля: ГКвозбуждается,
2)при освещении периферии поля: ГК тормозится
ганглиозные клетки с оff-рецептивным полем:
1) при освещении центра рецептивного поля: ГК тормозится,
2) при освещении периферии поля ГК возбуждается
Особенности рецептивных полей:
1. Рецептивные поля лучше реагируют на:
- движущиеся объекты,
- изменения освещённости,
- на контуры объектов
2. Все рецептивные поля – круглые .
3. Чем меньше рецептивные поля → тем лучше детализация изображения.
4. Диаметр рецептивного поля совпадает с протяжённостью дендритов ГК.
5. Рецептивные поля могут перекрываться. Это увеличивает надёжность восприятия сигнала .

П роводниковый
отдел зрительной
сенсорной системы:
роводниковый
отдел зрительной
сенсорной системы:
1. Зрительный нерв (аксоны ганглиозных клеток):
- Медиальные волокна делают перекрест
-Латеральные – нет
2. Зрительный тракт (после хиазмы)
3. Латеральные коленчатые тела таламуса (3-й чувствительный нейрон на пути переключения зрительной информации)
4. Зрительная лучистость
5. Стриарная кора затылочной доли
Центральный отдел зрительной сенсорной системы
Кора больших полушарий (затылочная доля)
Поля (Бродман):
- 17-первичное (стриарная кора)
- 18 – вторичное
- 19 – третичное
Неполный перекрест информации в зрительных нервах, позволяет анализировать в одном полушарии зрительную информацию от левого и правого глаза, для их сопоставления и измерения глубины полученного изображения.

Использование сразу двух глаз – бинокулярное зрение – дает возможность компенсировать повреждения одного глаза за счет другого, снимает эффект слепого пятна и лежит в основе стереоскопического зрения, когда на сетчатках одновременно возникают слегка различающиеся изображения одного и того же предмета, которые мозг воспринимает как один трехмерный образ. У человека общее поле зрения охватывает 180°, а стереоскопическое – 140°. Бинокулярость может нарушаться при косоглазии и некоторых других заболеваниях глаз. При сильной усталости может наблюдаться временное косоглазие, вызванное отключением ведомого глаза.
Как человек видит предметы объемными и как мы воспринимаем глубину пространства? Изображение окружающего мира на сетчатке является плоским. Однако мы видим мир объемным. Рассматривая предмет обоими глазами, мы видим его только тогда одиночным, когда оси зрения глаз образуют такой угол сходимости (конвергенцию), при котором симметричные отчетливые изображения на сетчатках получаются в определенных соответственных местах чувствительного желтого пятна (fovea centralis). Благодаря такому бинокулярному зрению, мы не только судим об относительном положении и расстоянии предметов, но и воспринимаем рельеф и объем. Существует несколько механизмов, которые обеспечивают построение объемного изображения пространства на основании плоских изображений, сформированных на сетчатке.
Во-первых, глаза расположены на некотором расстоянии друг от друга, поэтому изображения, формирующиеся на сетчатке левого и правого глаза, несколько различаются друг от друга. Чем ближе расположен объект по отношению к наблюдателю, тем больше будут различаться эти изображения.
Во-вторых, чем дальше расположен объект, тем меньше угловой размер его изображения на сетчатке. Оценивая угловые размеры знакомых предметов, мы можем оценивать расстояние до них.
В-третьих, перекрытие изображений также помогает оценить их взаимное расположение в пространстве. Изображение близкого предмета может перекрывать изображение удаленного, но не наоборот.
В-четвертых, при смещении головы наблюдателя изображения наблюдаемых объектов на сетчатке также будут смещаться (явление параллакса). При одном и том же смещении головы изображения близких объектов будут смещаться сильнее, чем изображения удаленных.
Преломляющие среды глаза: роговица, передняя камера, хрусталик, глазное яблоко
Примерно 2/3 из общей преломляющей силы глаза приходится на долю передней поверхности роговицы (не хрусталика глаза).
Общая преломляющая сила хрусталика глаза, когда он окружен со всех сторон жидкостью, составляет только 19 D (из общей - 59 D), то есть на его долю приходится примерно 1/3 от общей преломляющей силы глаза. Но значение хрусталика в том, что под влиянием нервной регуляции его кривизна может значительно изменяться, обеспечивая аккомодацию глаза – преломляющая сила хрусталика-33 D (из общей - 73 D)
Физическая рефракция. Рефракция глаза как физическое явление определяется радиусом кривизны каждой преломляющей среды глаза, показателями преломления сред и расстоянием между их поверхностями, то есть она обусловлена анатомическими особенностями глаза. Преломляющая сила может варьировать в широких пределах 52.0–68.0 D.
В жизни имеет значение не абсолютная сила оптического (светопреломляющего) аппарата глаза, а ее соотношение с длиной переднезадней оси глаза, то есть положение заднего главного фокуса (точка пересечения лучей, проходящих через оптическую систему глаза, параллельно его оптической оси) по отношению к сетчатке. Это и составляет понятие клинической рефракции.
Различают три вида клинической рефракции глаза:
-эмметропию
-миопию или близорукость
-гиперметропия или дальнозоркость.

Эмметропический глаз -преломляющая сила его оптической системы соответствует длине его оси, фокус параллельных лучей совпадает точно с сетчаткой, и такой глаз хорошо видит вдаль. Для зрения вблизи такому глазу необходимо усиливать свою рефракцию, что может быть достигнуто с помощью аккомодации.
Близорукость (миопия). Лучи собираются перед сетчаткой.
Причины:
· Анатомическая: длинная ось (27-28мм) глазного яблока (осевая близорукость)
· Функциональная: неспособность хрусталика расслабляться и становиться более плоским - цилиарная мышца не расслабляется (спазм аккомодации) – при длительном напряжении глаз.
Коррекция миопии: рассеивающие (двояковогнутые) линзы
Например: длина глаза 26 мм, преломляющая сила 67 Д. Рассеивающая линза «-8» «мешает» преломлению: уменьшает его на 8 Д: 67-8=59 Д.
Дальнозоркость. Лучи собираются за сетчаткой
Причины:
· Анатомическая: короткое глазное яблоко (гиперметропия). Дети рождаются дальнозоркими (недоразвито глазное яблоко). До 7 лет дальнозоркость – норма.
· Функциональная: неспособность хрусталика аккомодировать и становиться более выпуклым (пресбиопия, или старческая дальнозоркость):
- Утрата эластичности
- Слабая цилиарная мышца
Коррекция дальнозоркости - собирающие (двояковыпуклые) линзы
Например: длина глаза 20 мм, преломляющая сила 55 Д. Собирающая линза «+4» «помогает»: увеличивает преломление ещё на 4 Д: 55+4=59 Д.
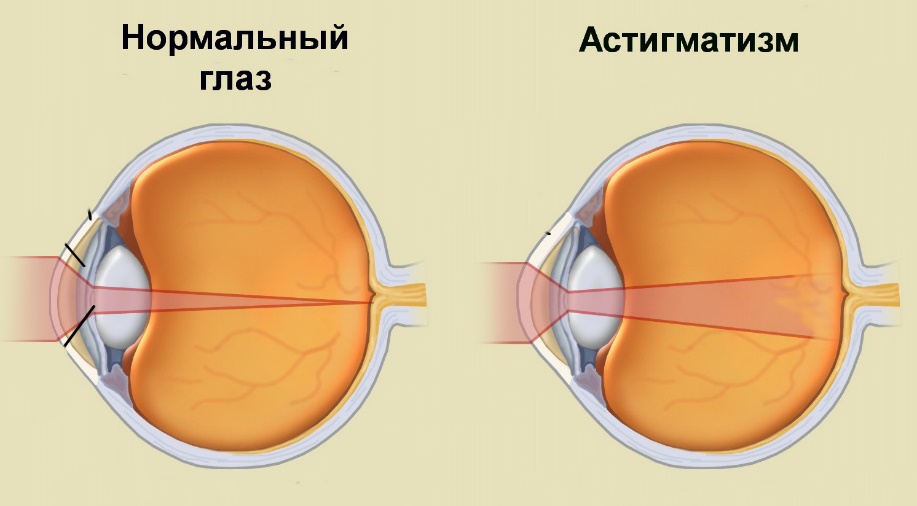
Астигматизм:
В норме разница разница преломления по вертикали и горизонтали не более 0,5 Д. При чрезмерной выраженности – астигматизм. Световой пучок невозможно собрать в одной точке, возникает несколько фокусов. Происходит искажение преломления из-за неодинаковой кривизны роговицы и/или хрусталика в разных меридианах - искажение высоты и ширины предметов, раздвоение контуров предметов.
Коррекция астигматизма – сложные цилиндрические линзы. В одном стекле: несколько линз (цилиндров) с разными преломляющими силами

Слуховая сенсорная система
Слуховой анализатор включает в себя:
- периферический отдел, воспринимающий звуковые колебания (колебания воздуха или другой упругой среды) и передающих их на слуховые рецепторы,
- проводниковой
- корковой части. Особая роль слухового анализатора у человека связана с членораздельной речью.
Периферический – рецепторный отдел
- это Кортиев орган – вторично-чувствующие рецепторы. Находятся в улитке внутреннего уха, которая расположена в пирамиде височной кости. Звуковые колебания передаются к ним через систему вспомогательных образований наружного, среднего и внутреннего уха.

Наружное ухо – ушная раковина и слуховой проход – служат для улавливания звуков и усиления звуковых колебаний. Ухо человека воспринимает колебания с частотой от 16 до 20 тысяч Герц. На границе наружного и среднего уха располагается барабанная перепонка, которая колеблется при действии звуковых колебаний.
Среднее ухо состоит из барабанной полости, в которой расположены слуховые косточки:
- молоточек,
- наковальня,
- стремечко,
по которым колебания барабанной перепонки передаются на перепонку овального окна, отделяющего среднее ухо от улитки внутреннего уха. Площадь барабанной перепонки значительно больше площади овального окна, что приводит к усилению колебаний в 45 раз.
Барабанный рефлекс – механизм ослабления звука, срабатывает при громких звуках за счёт расслабления мышцы, натягивающей барабанную перепонку (m. tympani), и сокращения мышцы, фиксирующей стремечко (m. stapedius). Но латентный период барабанного рефлекса 40-160 мс. Поэтому он не может защитить от коротких интенсивных неожиданных звуков (выстрел, взрыв), в результате может произойти разрыв барабанной перепонки. Эти же мышцы сокращаются не только при действии посторонних громких звуков, но и во время издавания звуков самим человеком (речь, крик). Это так называемый предвокализационный рефлекс. Поэтому окружающий шум нам кажется меньше, если мы сами в это время громко говорим ("перекрикиваем" шум).
В полости среднего уха (барабанной полости) находится воздух, давление которого равно атмосферному за счет слуховой трубы, соединяющей барабанную полость с глоткой. При глотании слуховая труба открывается, и давление в среднем ухе выравнивается с атмосферным.
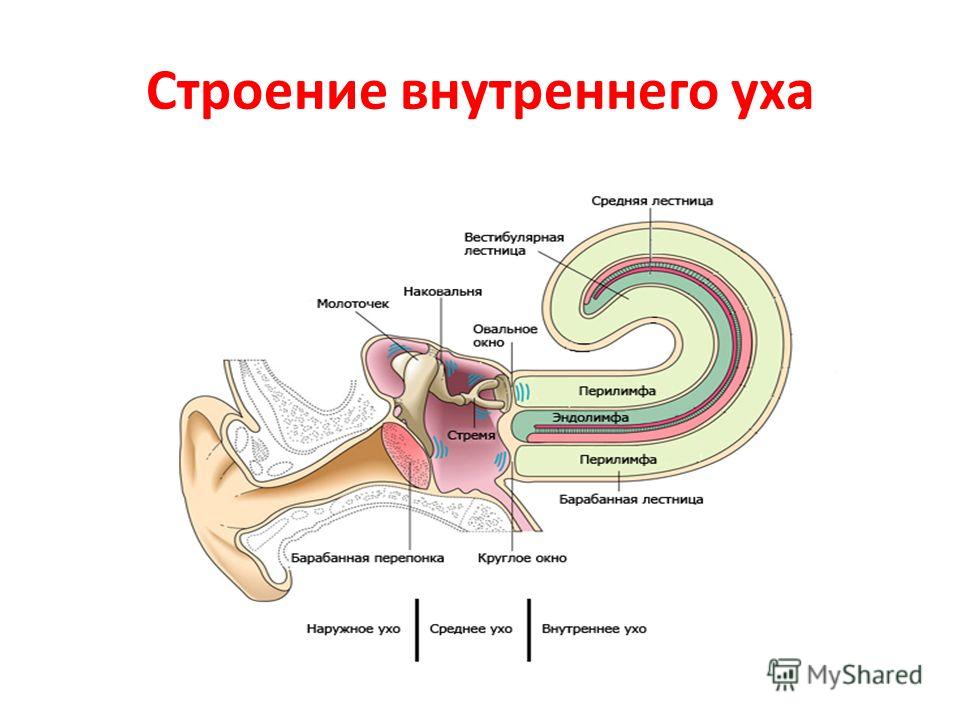
Внутреннее ухо располагается в пирамиде височной кости.
Костная улитка – спирально извивающийся канал, образует 2,5 оборота вокруг стержня. Внутри канала улитки идут 2 мембраны – рейснерова и базилярная, которые образуют 3 «лестницы»:
- вестибулярную,
- среднюю (улитковый ход)
- барабанную.
В вестибулярной и барабанной лестницах находится перилимфа (по составу солей похожа на внеклеточную жидкость, ликвор), в средней (улитковом ходе) – эндолимфа (содержит в 100 раз больше К+ и в 10 раз меньше Na+ - как в цитоплазме клеток). Это способствует образованию рецепторного потенциала и повышает чувствительность слуховых рецепторов к колебаниям эндолимфы. Вестибулярная и барабанная лестницы соединяются между собой с помощью отверстия геликотремы.
Колебания стремечка передаются через мембрану овального окна на перилимфу вестибулярной лестницы и через геликотрему на перилимфу барабанной лестницы. Эти колебания передаются на перепонку круглого окна и одновременно заставляют колебаться эндолимфу и базилярную мембрану.
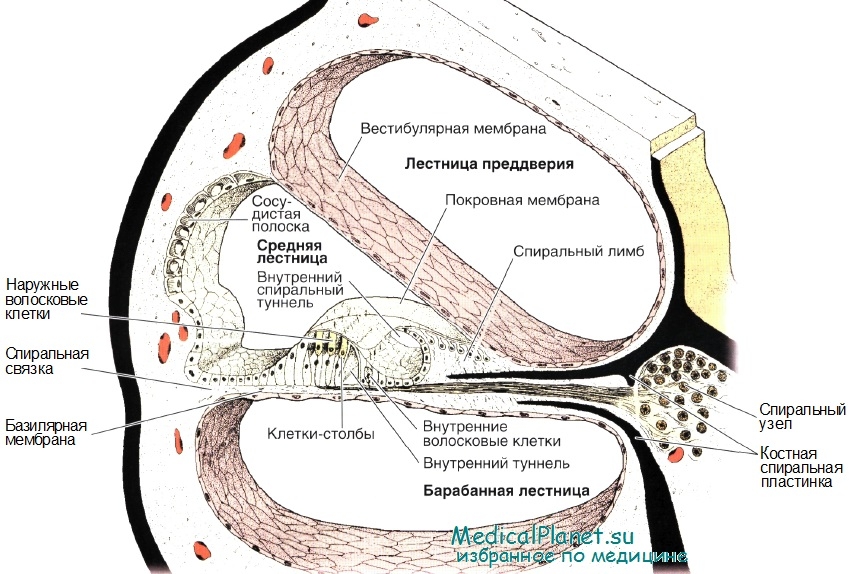
В улитковом ходе на базилярной мембране располагаются рецепторные слуховые клетки, которые вместе с базилярной мембраной составляют Кортиев орган (периферический отдел слухового анализатора). Рецепторные слуховые клетки кортиева органа имеют волоски (цилии). Над кортиевым органом расположена перепонка – покровная, текториальная или кортиева, мембрана.

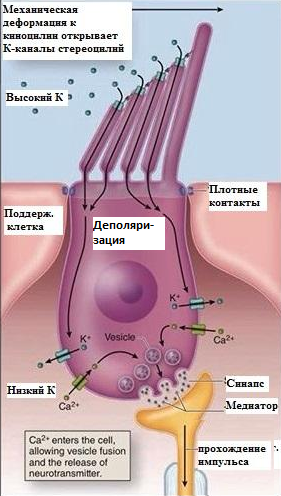
Звуковые колебания воспринимаются рецепторными волосковыми клетками Кортиева органа.
Мембранный потенциал покоя слуховой рецепторной клетки равен –60 мВ.
В результате деформации волосков, открываются механочувствительные калиевые каналы на мембране → ток калия внутрь рецепторной клетки (пассивно, по градиенту концентрации из эндолимфы )→ рецепторный потенциал (деполяризационный электротонический) → выброс медиатора – глутамата → возбуждение дендритов 1 чувствительного нейрона (биполярной клетки, тело которой лежит в спиральном ганглии рядом с улиткой), генерация на е аксональном холмикеПД и проведение ПД (аксоны биполярной клетки формируют слуховой нерв).
Т.о. РП волосковой клетки деполяризационный, связанный с током внутрь клетки К+.